

High resolution approaches for the identification of amyloid fragments in brain
- PMID: 30367888
- PMCID: PMC6451891
- DOI: 10.1016/j.jneumeth.2018.10.032
High resolution approaches for the identification of amyloid fragments in brain
Abstract
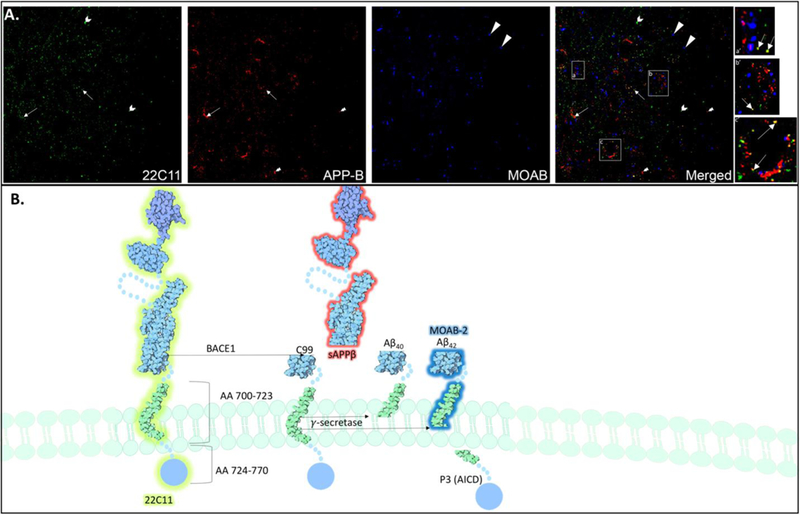
Background: It is now widely recognized that endogenous, picomolar concentrations of the 42 amino acid long peptide, amyloid-β (Aβ42) is secreted under normal physiological conditions and exerts important functional activity throughout neuronal intracellular compartments. Transgenic animal models that overexpress Aβ42 and its precursor, amyloid precursor protein (APP), have not provided predictive value in testing new treatments for Alzheimer's disease (AD), resulting in failed clinical trials. While these results are discouraging, they underscore the need to understand the physiological roles of Aβ42 and APP under normal conditions as well as at early pre- symptomatic stages of AD. New method: We describe the use of acrolein-perfusion in immunoelectron microscopy in combination with novel antibodies directed against endogenous murine Aβ42 and APP fragments to study abnormalities in the endolysosomal system at early stages of disease. The specific requirements, limitations and advantages of novel antibodies directed against human and murine Aβ42, APP and APP fragments are discussed as well as parameters for ultrastructural analysis of endolysosomal compartments.
Results: Novel antibodies and a detailed protocol for immunoelectron microscopy using acrolein as a fixative are described. Acrolein is shown to preserve intraneuronal Aβ42 species, as opposed to paraformaldehyde fixed tissue, which primarily preserves membrane bound species. Comparison with existing method(s): Technology sensitive enough to detect endogenous Aβ42 under physiological conditions has not been widely available. We describe a number of novel and highly sensitive antibodies have recently been developed that may facilitate the analysis of endogenous Aβ42.
Conclusions: Using novel and highly specific antibodies in combination with electron microscopy may reveal important information about the timing of aberrant protein accumulation, as well as the progression of abnormalities in the endolysosomal systems that sort and clear these peptides.
Keywords: Amyloid; Antibodies; Electron microscopy.
Copyright © 2018. Published by Elsevier B.V.
Figures




References
-
- Barnham KJ, McKinstry WJ, Multhaup G, Galatis D, Morton CJ, Curtain CC, Williamson NA, White AR, Hinds MG, Norton RS, Beyreuther K, Masters CL, Parker MW and Cappai R (2003). “Structure of the Alzheimer’s disease amyloid precursor protein copper binding domain. A regulator of neuronal copper homeostasis.” J Biol Chem 278(19): 17401–17407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
