

Species Widely Distributed in Halophilic Archaea Exhibit Opsin-Mediated Inhibition of Bacterioruberin Biosynthesis
- PMID: 30373756
- PMCID: PMC6304663
- DOI: 10.1128/JB.00576-18
Species Widely Distributed in Halophilic Archaea Exhibit Opsin-Mediated Inhibition of Bacterioruberin Biosynthesis
Abstract
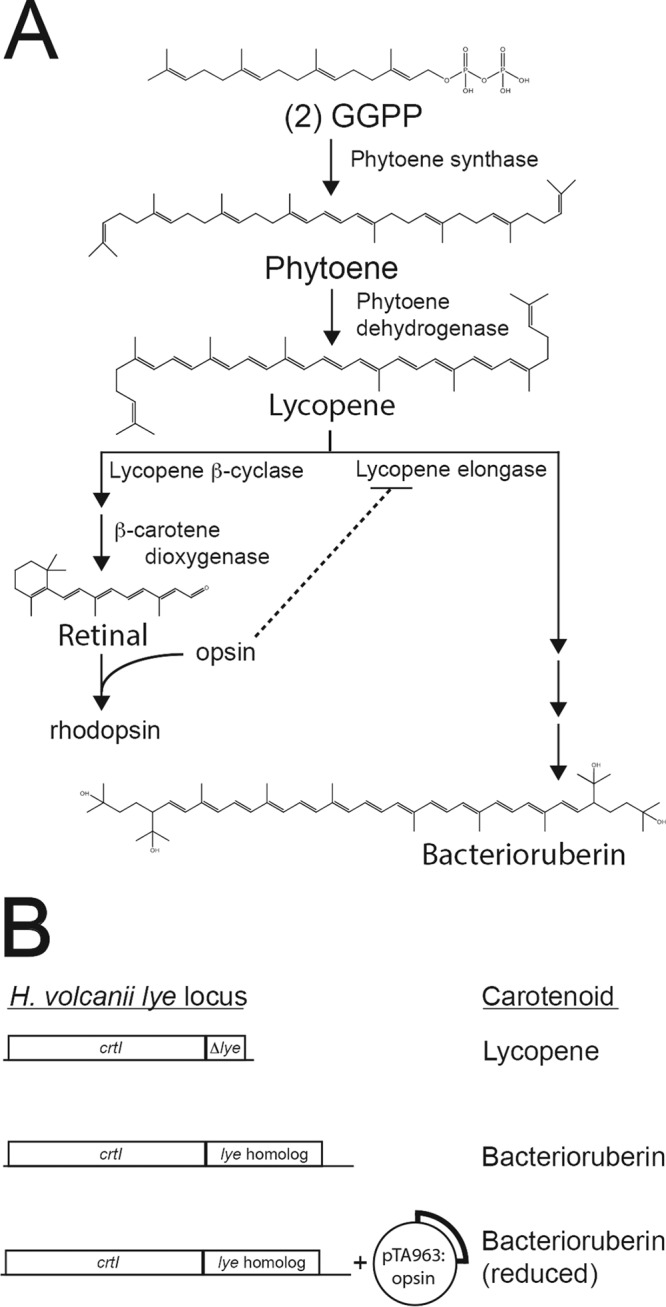
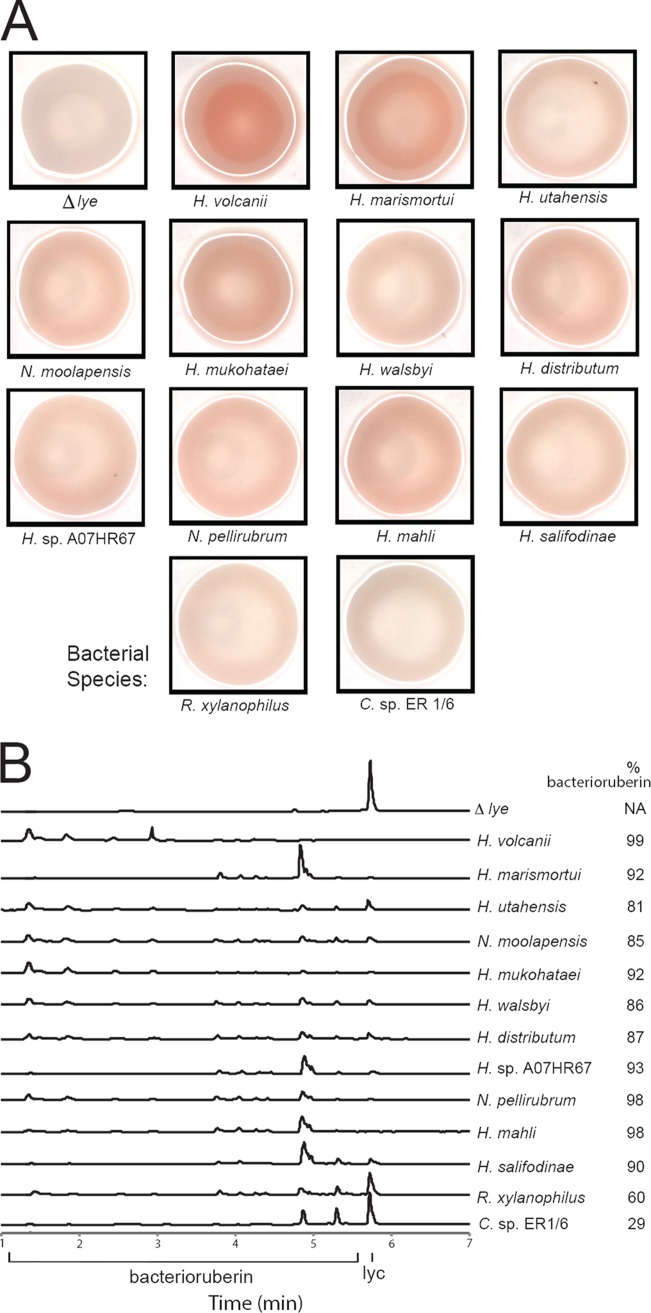
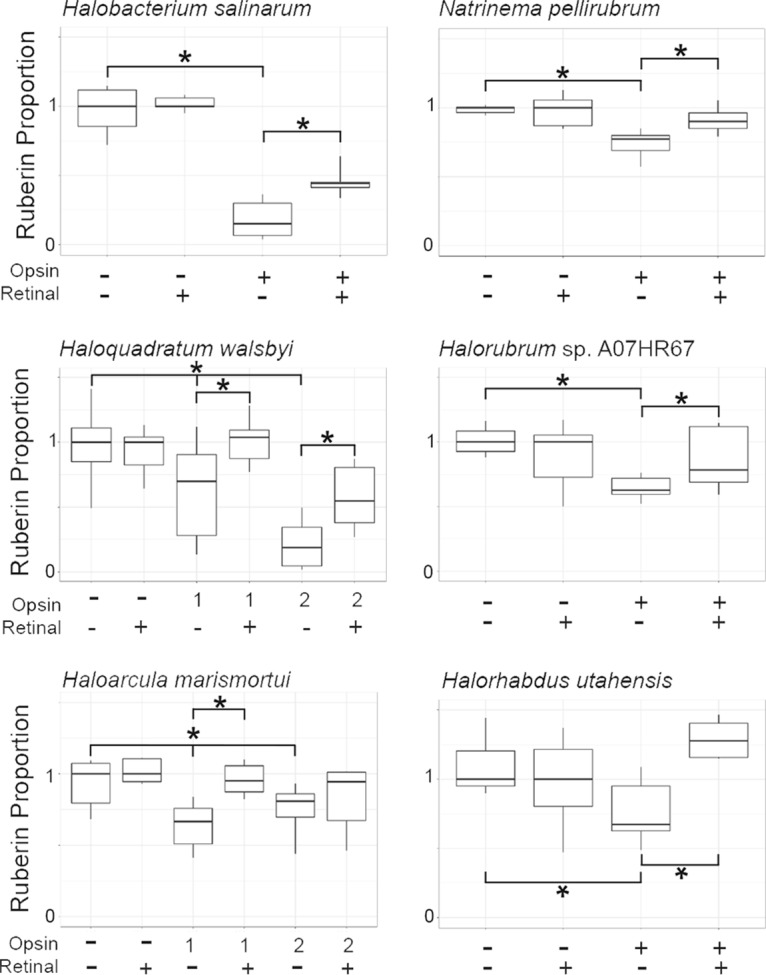
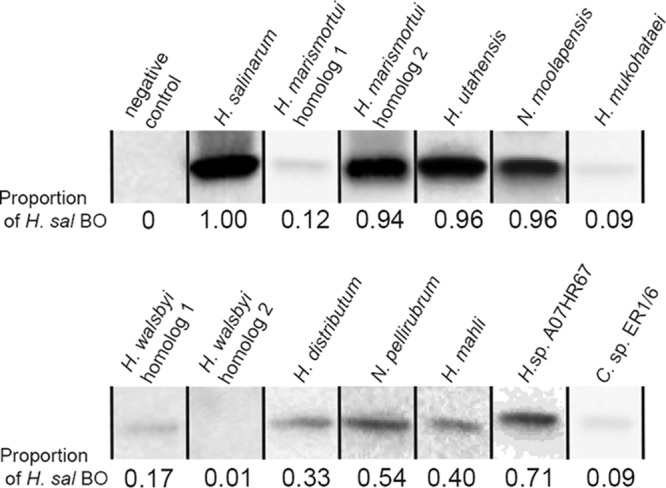
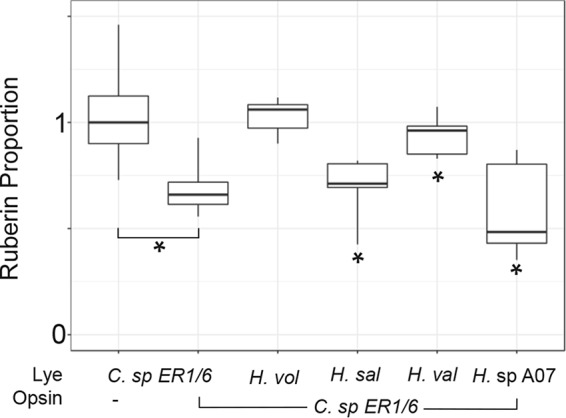
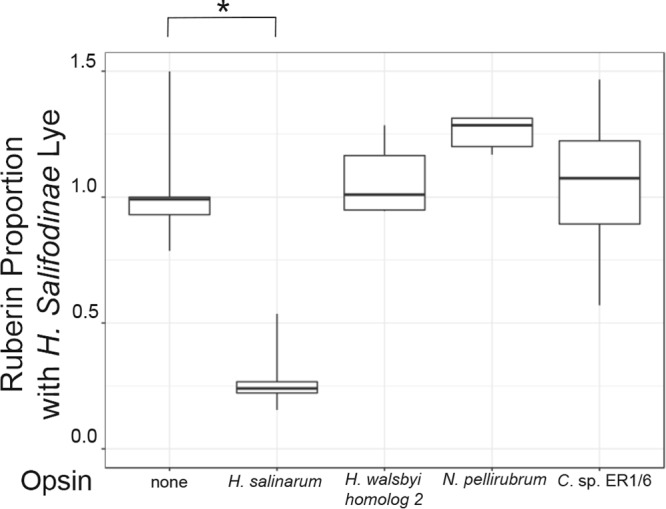
Halophilic Archaea are a distinctive pink color due to a carotenoid pigment called bacterioruberin. To sense or utilize light, many halophilic Archaea also produce rhodopsins, complexes of opsin proteins with a retinal prosthetic group. Both bacterioruberin and retinal are synthesized from isoprenoid precursors, with lycopene as the last shared intermediate. We previously described a regulatory mechanism by which Halobacterium salinarum bacterioopsin and Haloarcula vallismortis cruxopsin inhibit bacterioruberin synthesis catalyzed by lycopene elongase. In this work, we found that opsins in all three major Halobacteria clades inhibit bacterioruberin synthesis, suggesting that this regulatory mechanism existed in the common Halobacteria ancestor. Halophilic Archaea, which are generally heterotrophic and aerobic, likely evolved from an autotrophic, anaerobic methanogenic ancestor by acquiring many genes from Bacteria via lateral gene transfer. These bacterial "imports" include genes encoding opsins and lycopene elongases. To determine if opsins from Bacteria inhibit bacterioruberin synthesis, we tested bacterial opsins and found that an opsin from Curtobacterium, in the Actinobacteria phylum, inhibits bacterioruberin synthesis catalyzed by its own lycopene elongase, as well as that catalyzed by several archaeal enzymes. We also determined that the lycopene elongase from Halococcus salifodinae, a species from a family of Halobacteria lacking opsin homologs, retained the capacity to be inhibited by opsins. Together, our results indicate that opsin-mediated inhibition of bacterioruberin biosynthesis is a widely distributed mechanism found in both Archaea and Bacteria, possibly predating the divergence of the two domains. Further analysis may provide insight into the acquisition and evolution of the genes and their host species.IMPORTANCE All organisms use a variety of mechanisms to allocate limited resources to match their needs in their current environment. Here, we explore how halophilic microbes use a novel mechanism to allow efficient production of rhodopsin, a complex of an opsin protein and a retinal prosthetic group. We previously demonstrated that Halobacterium salinarum bacterioopsin directs available resources toward retinal by inhibiting synthesis of bacterioruberin, a molecule that shares precursors with retinal. In this work, we show that this mechanism can be carried out by proteins from halophilic Archaea that are not closely related to H. salinarum and those in at least one species of Bacteria Therefore, opsin-mediated inhibition of bacterioruberin synthesis may be a highly conserved, ancient regulatory mechanism.
Keywords: C50 carotenoid; UbiA prenyltransferase; carotenoid biosynthesis; cofactor biosynthesis; membrane protein biogenesis; microbial rhodopsin; proteorhodopsin.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Opsin-Mediated Inhibition of Bacterioruberin Synthesis in Halophilic Archaea.J Bacteriol. 2017 Oct 3;199(21):e00303-17. doi: 10.1128/JB.00303-17. Print 2017 Nov 1. J Bacteriol. 2017. PMID: 28784816 Free PMC article.
-
Bacterioopsin-mediated regulation of bacterioruberin biosynthesis in Halobacterium salinarum.J Bacteriol. 2011 Oct;193(20):5658-67. doi: 10.1128/JB.05376-11. Epub 2011 Aug 12. J Bacteriol. 2011. PMID: 21840984 Free PMC article.
-
Complete biosynthetic pathway of the C50 carotenoid bacterioruberin from lycopene in the extremely halophilic archaeon Haloarcula japonica.J Bacteriol. 2015 May;197(9):1614-23. doi: 10.1128/JB.02523-14. Epub 2015 Feb 23. J Bacteriol. 2015. PMID: 25712483 Free PMC article.
-
Carotenoid Production by Halophilic Archaea Under Different Culture Conditions.Curr Microbiol. 2016 May;72(5):641-51. doi: 10.1007/s00284-015-0974-8. Epub 2016 Jan 11. Curr Microbiol. 2016. PMID: 26750123 Review.
-
The information transfer system of halophilic archaea.Plasmid. 2011 Mar;65(2):77-101. doi: 10.1016/j.plasmid.2010.11.005. Epub 2010 Nov 19. Plasmid. 2011. PMID: 21094181 Review.
Cited by
-
Proteolysis at the Archaeal Membrane: Advances on the Biological Function and Natural Targets of Membrane-Localized Proteases in Haloferax volcanii.Front Microbiol. 2022 Jun 24;13:940865. doi: 10.3389/fmicb.2022.940865. eCollection 2022. Front Microbiol. 2022. PMID: 35814708 Free PMC article. Review.
-
The Blue-Green Sensory Rhodopsin SRM from Haloarcula marismortui Attenuates Both Phototactic Responses Mediated by Sensory Rhodopsin I and II in Halobacterium salinarum.Sci Rep. 2019 Apr 5;9(1):5672. doi: 10.1038/s41598-019-42193-y. Sci Rep. 2019. PMID: 30952934 Free PMC article.
-
Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico.Microorganisms. 2021 May 20;9(5):1096. doi: 10.3390/microorganisms9051096. Microorganisms. 2021. PMID: 34065163 Free PMC article.
-
Cellular differentiation into hyphae and spores in halophilic archaea.Nat Commun. 2023 Apr 1;14(1):1827. doi: 10.1038/s41467-023-37389-w. Nat Commun. 2023. PMID: 37005419 Free PMC article.
References
-
- Lazrak T, Wolff G, Albrecht A-M, Nakatani Y, Ourisson G, Kates M. 1988. Bacterioruberins reinforce reconstituted Halobacterium lipid membranes. Biochim Biophys Acta 939:160–162. doi:10.1016/0005-2736(88)90057-0. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
