

Regulatory discrimination of mRNAs by FMRP controls mouse adult neural stem cell differentiation
- PMID: 30373821
- PMCID: PMC6275535
- DOI: 10.1073/pnas.1809588115
Regulatory discrimination of mRNAs by FMRP controls mouse adult neural stem cell differentiation
Abstract
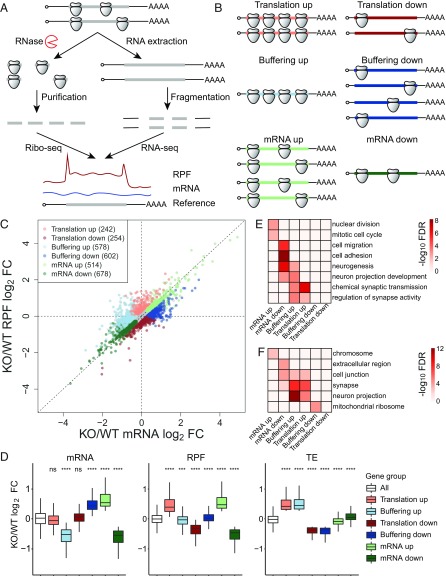
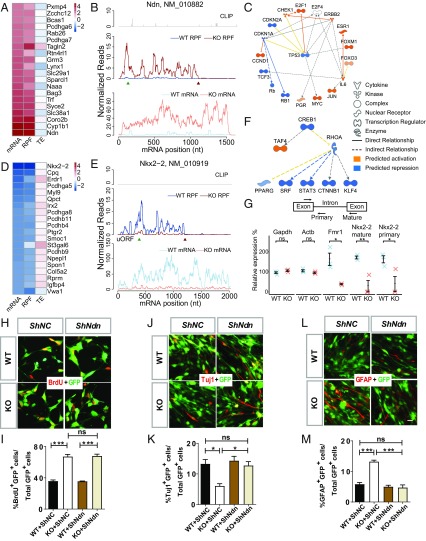
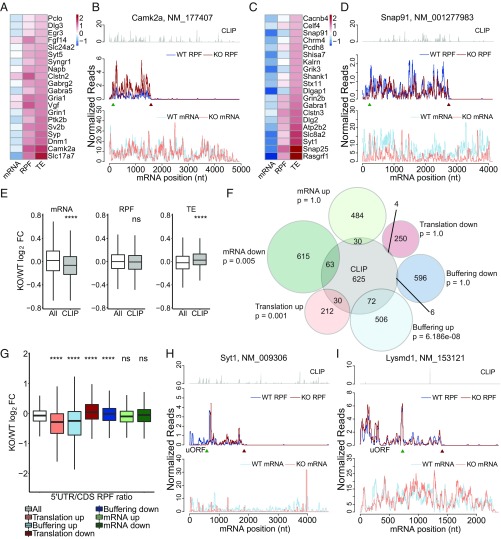
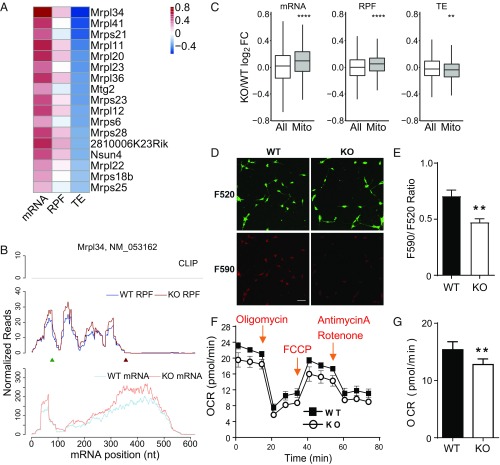
Fragile X syndrome (FXS) is caused by the loss of fragile X mental retardation protein (FMRP), an RNA binding protein whose deficiency impacts many brain functions, including differentiation of adult neural stem cells (aNSCs). However, the mechanism by which FMRP influences these processes remains unclear. Here, we performed ribosome profiling and transcriptomic analysis of aNSCs in parallel from wild-type and Fmr1 knockout mice. Our data revealed diverse gene expression changes at both mRNA and translation levels. Many mitosis and neurogenesis genes were dysregulated primarily at the mRNA level, while numerous synaptic genes were mostly dysregulated at the translation level. Translational "buffering", whereby changes in ribosome association with mRNA are compensated by alterations in RNA abundance, was also evident. Knockdown of NECDIN, an FMRP-repressed transcriptional factor, rescued neuronal differentiation. In addition, we discovered that FMRP regulates mitochondrial mRNA expression and energy homeostasis. Thus, FMRP controls diverse transcriptional and posttranscriptional gene expression programs critical for neural differentiation.
Keywords: fragile X syndrome; neural differentiation; neural stem cells; ribosome profiling; translation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
"Fragile" equilibrium between translation and transcription.Proc Natl Acad Sci U S A. 2018 Nov 27;115(48):12086-12088. doi: 10.1073/pnas.1817562115. Epub 2018 Nov 14. Proc Natl Acad Sci U S A. 2018. PMID: 30429319 Free PMC article. No abstract available.
References
-
- Santoro MR, Bray SM, Warren ST. Molecular mechanisms of fragile X syndrome: A twenty-year perspective. Annu Rev Pathol. 2012;7:219–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
