

Engineering yeast endosymbionts as a step toward the evolution of mitochondria
- PMID: 30373839
- PMCID: PMC6243291
- DOI: 10.1073/pnas.1813143115
Engineering yeast endosymbionts as a step toward the evolution of mitochondria
Abstract
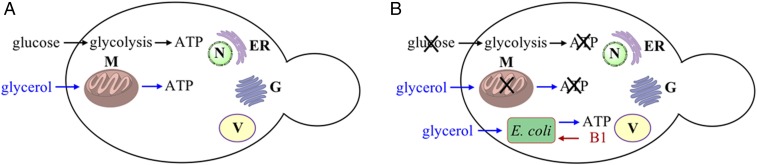
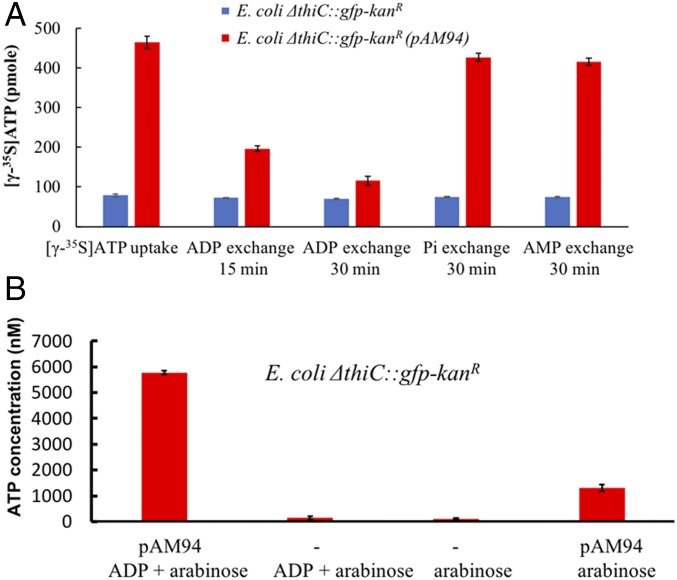
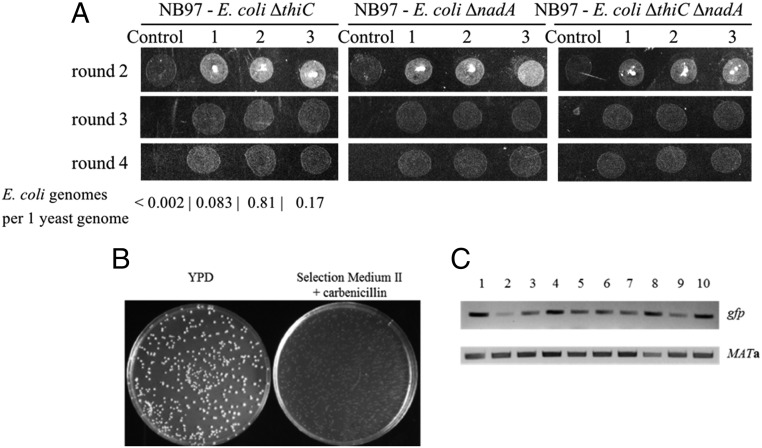
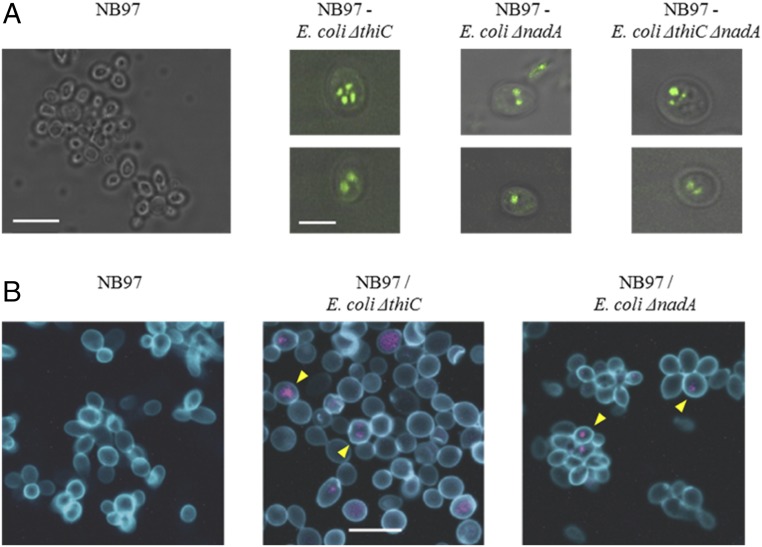
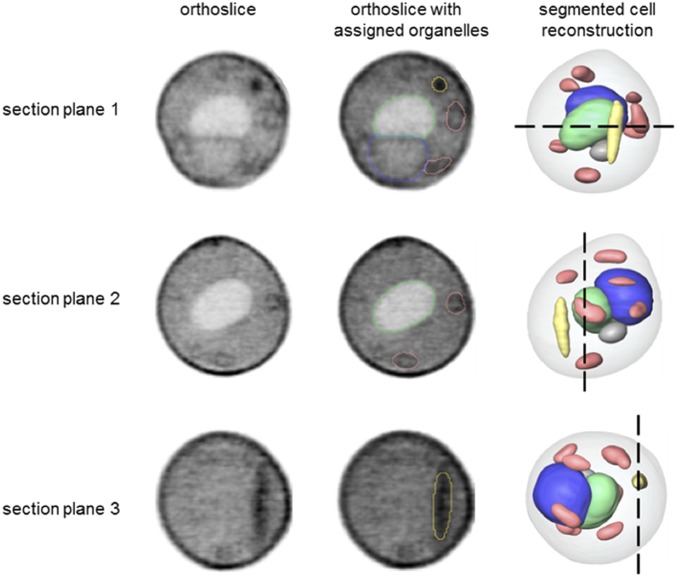
It has been hypothesized that mitochondria evolved from a bacterial ancestor that initially became established in an archaeal host cell as an endosymbiont. Here we model this first stage of mitochondrial evolution by engineering endosymbiosis between Escherichia coli and Saccharomyces cerevisiae An ADP/ATP translocase-expressing E. coli provided ATP to a respiration-deficient cox2 yeast mutant and enabled growth of a yeast-E. coli chimera on a nonfermentable carbon source. In a reciprocal fashion, yeast provided thiamin to an endosymbiotic E. coli thiamin auxotroph. Expression of several SNARE-like proteins in E. coli was also required, likely to block lysosomal degradation of intracellular bacteria. This chimeric system was stable for more than 40 doublings, and GFP-expressing E. coli endosymbionts could be observed in the yeast by fluorescence microscopy and X-ray tomography. This readily manipulated system should allow experimental delineation of host-endosymbiont adaptations that occurred during evolution of the current, highly reduced mitochondrial genome.
Keywords: ADP/ATP translocase; endosymbiotic theory; evolution; mitochondria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Margulis L. Origin of Eukaryotic Cells: Evidence and Research Implications for a Theory of the Origin and Evolution of Microbial, Plant and Animal Cells on the Precambrian Earth. Yale Univ Press; New Haven, CT: 1970.
-
- Kutschera U, Niklas KJ. The modern theory of biological evolution: An expanded synthesis. Naturwissenschaften. 2004;91:255–276. - PubMed
-
- Archibald JM. Endosymbiosis and eukaryotic cell evolution. Curr Biol. 2015;25:R911–R921. - PubMed
-
- Javaux EJ, Knoll AH, Walter MR. Morphological and ecological complexity in early eukaryotic ecosystems. Nature. 2001;412:66–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
