

Organellar TRP channels
- PMID: 30374082
- PMCID: PMC6785982
- DOI: 10.1038/s41594-018-0148-z
Organellar TRP channels
Abstract
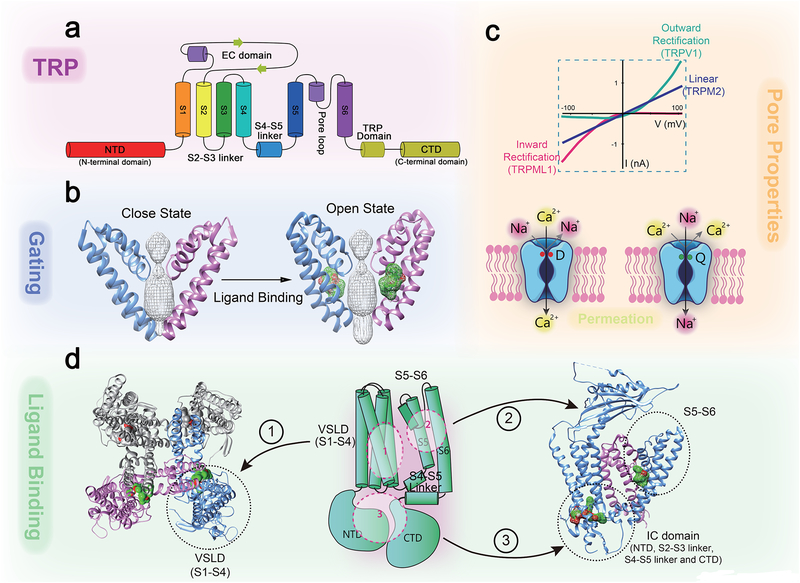
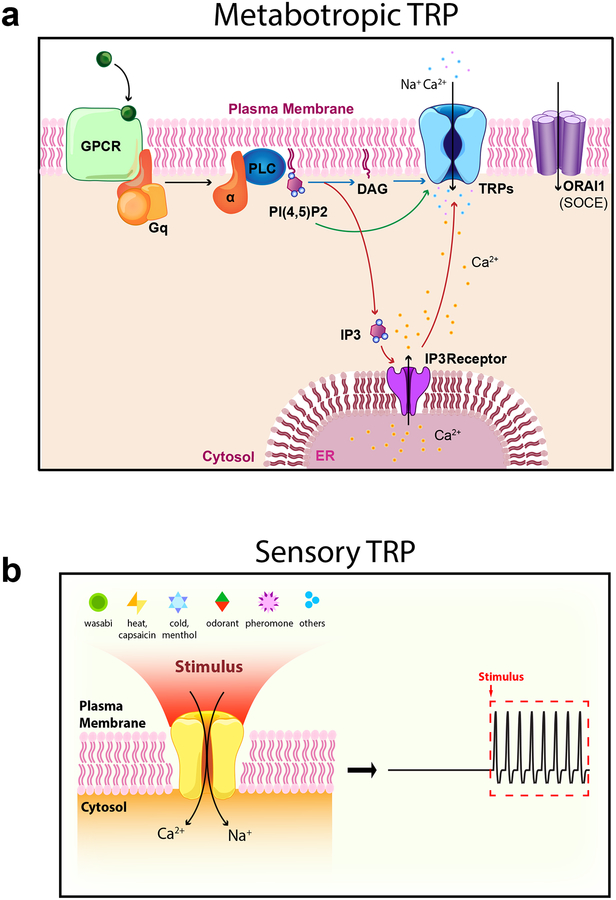
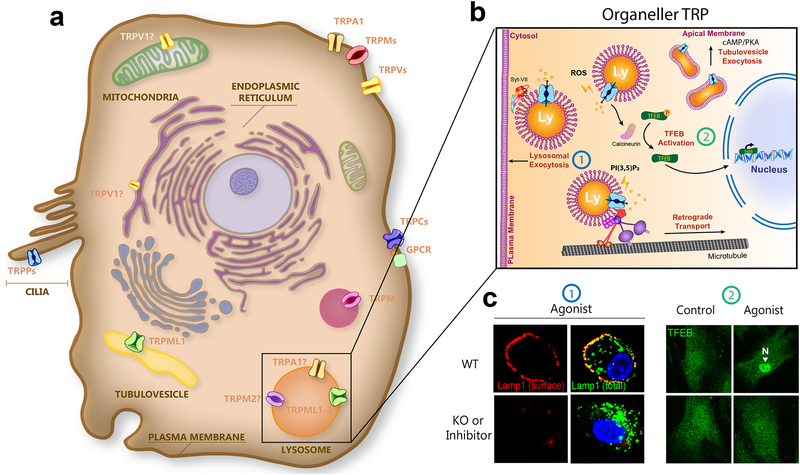
Mammalian transient receptor potential (TRP) channels mediate Ca2+ flux and voltage changes across membranes in response to environmental and cellular signals. At the plasma membrane, sensory TRPs act as neuronal detectors of physical and chemical environmental signals, and receptor-operated (metabotropic) TRPs decode extracellular neuroendocrine cues to control body homeostasis. In intracellular membranes, such as those in lysosomes, organellar TRPs respond to compartment-derived signals to control membrane trafficking, signal transduction, and organelle function. Complementing mouse and human genetics and high-resolution structural approaches, physiological studies employing natural agonists and synthetic inhibitors have become critical in resolving the in vivo functions of metabotropic, sensory, and organellar TRPs.
Figures

References
-
- Minke B, Wu C & Pak WL Induction of photoreceptor voltage noise in the dark in Drosophila mutant. Nature 258, 84–87 (1975). - PubMed
-
- Montell C & Rubin GM Molecular characterization of the Drosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron 2, 1313–1323 (1989). - PubMed
-
- Hardie RC & Minke B The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron 8, 643–651 (1992). - PubMed
-
- Zhu X, Chu PB, Peyton M & Birnbaumer L Molecular cloning of a widely expressed human homologue for the Drosophila trp gene. FEBS Lett 373, 193–198 (1995). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
