

The multicatalytic compartment of propionyl-CoA synthase sequesters a toxic metabolite
- PMID: 30374166
- PMCID: PMC6499725
- DOI: 10.1038/s41589-018-0153-x
The multicatalytic compartment of propionyl-CoA synthase sequesters a toxic metabolite
Abstract
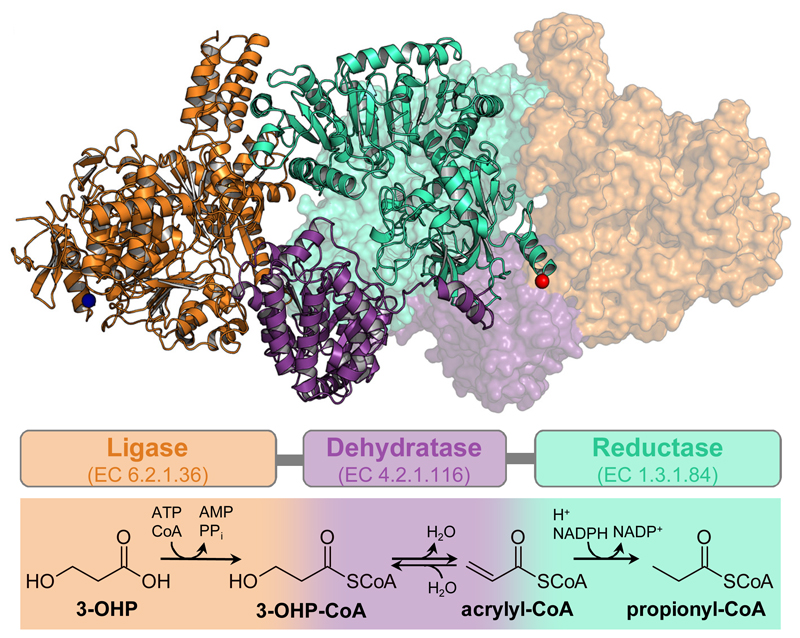
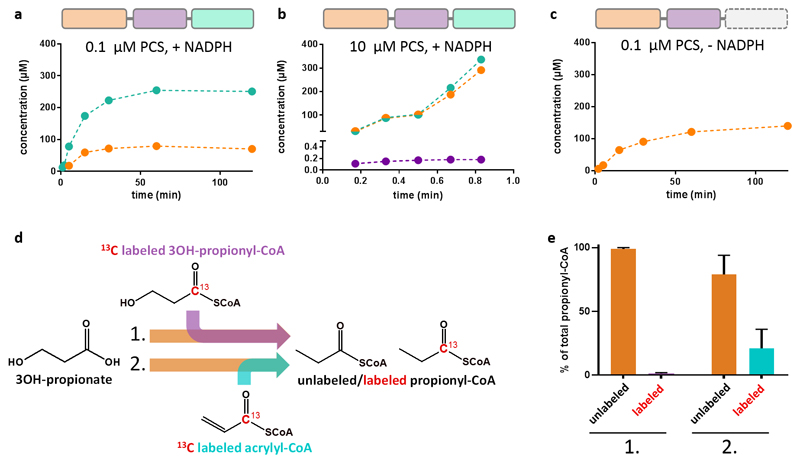
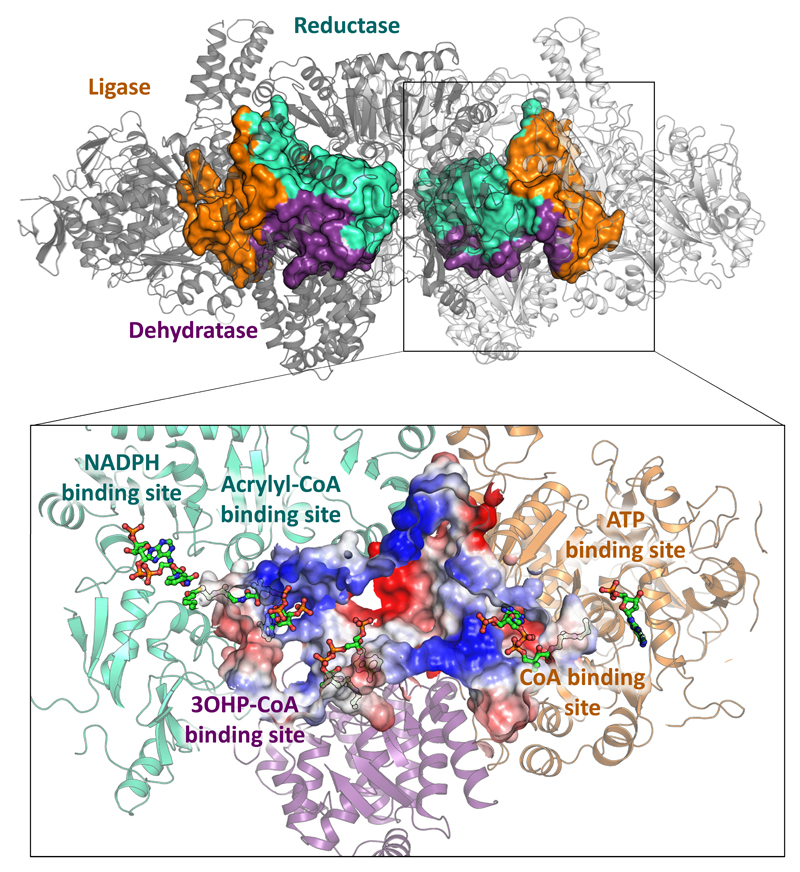
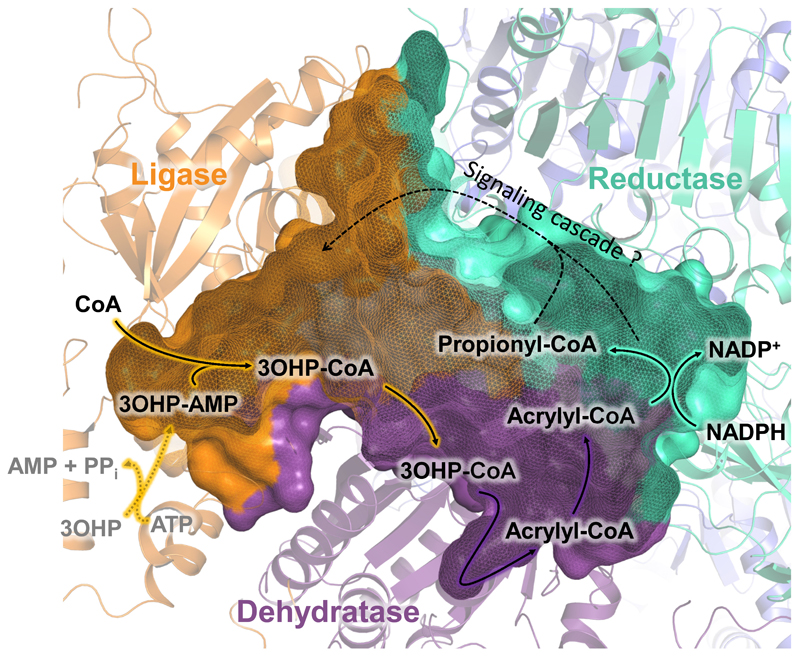
Cells must cope with toxic or reactive intermediates formed during metabolism. One coping strategy is to sequester reactions that produce such intermediates within specialized compartments or tunnels connecting different active sites. Here, we show that propionyl-CoA synthase (PCS), an ∼ 400-kDa homodimer, three-domain fusion protein and the key enzyme of the 3-hydroxypropionate bi-cycle for CO2 fixation, sequesters its reactive intermediate acrylyl-CoA. Structural analysis showed that PCS forms a multicatalytic reaction chamber. Kinetic analysis suggested that access to the reaction chamber and catalysis are synchronized by interdomain communication. The reaction chamber of PCS features three active sites and has a volume of only 33 nm3. As one of the smallest multireaction chambers described in biology, PCS may inspire the engineering of a new class of dynamically regulated nanoreactors.
Conflict of interest statement
Figures
References
-
- Wheeldon I, et al. Substrate channelling as an approach to cascade reactions. Nat Chem. 2016;8:299–309. - PubMed
-
- Linster CL, Van Schaftingen E, Hanson AD. Metabolite damage and its repair or pre-emption. Nat Chem Biol. 2013;9:72–80. - PubMed
-
- Wheeldon I, et al. Substrate channelling as an approach to cascade reactions. Nature chemistry. 2016;8:299. - PubMed
-
- Alber BE, Fuchs G. Propionyl-coenzyme A synthase from Chloroflexus aurantiacus, a key enzyme of the 3-hydroxypropionate cycle for autotrophic CO2 fixation. J Biol Chem. 2002;277:12137–12143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
