

Lon protease inactivation in Drosophila causes unfolded protein stress and inhibition of mitochondrial translation
- PMID: 30374414
- PMCID: PMC6197249
- DOI: 10.1038/s41420-018-0110-1
Lon protease inactivation in Drosophila causes unfolded protein stress and inhibition of mitochondrial translation
Erratum in
-
Erratum: Publisher Correction: articles initially published in wrong volume.Cell Death Discov. 2019 Jul 10;5:116. doi: 10.1038/s41420-019-0186-2. eCollection 2019. Cell Death Discov. 2019. PMID: 31312525 Free PMC article.
Abstract
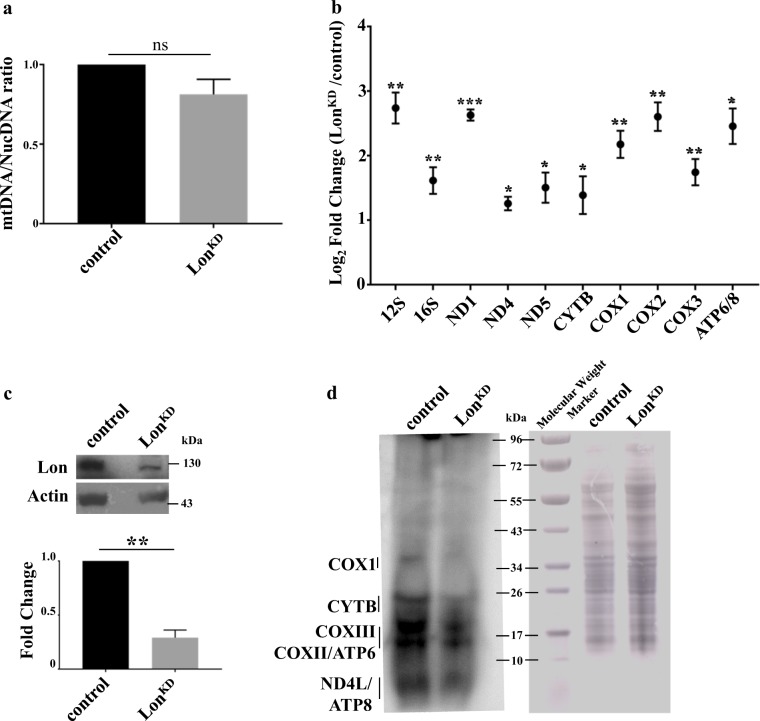
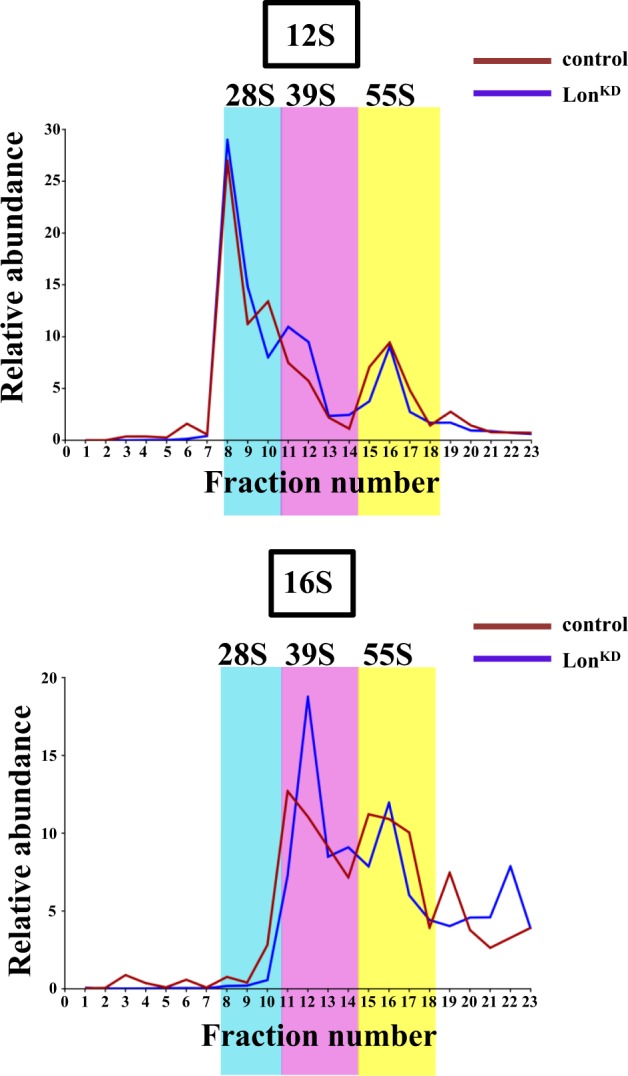
Mitochondrial dysfunction is a frequent participant in common diseases and a principal suspect in aging. To combat mitochondrial dysfunction, eukaryotes have evolved a large repertoire of quality control mechanisms. One such mechanism involves the selective degradation of damaged or misfolded mitochondrial proteins by mitochondrial resident proteases, including proteases of the ATPase Associated with diverse cellular Activities (AAA+) family. The importance of the AAA+ family of mitochondrial proteases is exemplified by the fact that mutations that impair their functions cause a variety of human diseases, yet our knowledge of the cellular responses to their inactivation is limited. To address this matter, we created and characterized flies with complete or partial inactivation of the Drosophila matrix-localized AAA+ protease Lon. We found that a Lon null allele confers early larval lethality and that severely reducing Lon expression using RNAi results in shortened lifespan, locomotor impairment, and respiratory defects specific to respiratory chain complexes that contain mitochondrially encoded subunits. The respiratory chain defects of Lon knockdown (Lon KD ) flies appeared to result from severely reduced translation of mitochondrially encoded genes. This translational defect was not a consequence of reduced mitochondrial transcription, as evidenced by the fact that mitochondrial transcripts were elevated in abundance in Lon KD flies. Rather, the translational defect of Lon KD flies appeared to be derived from sequestration of mitochondrially encoded transcripts in highly dense ribonucleoparticles. The translational defect of Lon KD flies was also accompanied by a substantial increase in unfolded mitochondrial proteins. Together, our findings suggest that the accumulation of unfolded mitochondrial proteins triggers a stress response that culminates in the inhibition of mitochondrial translation. Our work provides a foundation to explore the underlying molecular mechanisms.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures





Similar articles
-
Inactivation of Lon protease reveals a link between mitochondrial unfolded protein stress and mitochondrial translation inhibition.Cell Death Dis. 2018 Dec 5;9(12):1168. doi: 10.1038/s41419-018-1213-6. Cell Death Dis. 2018. PMID: 30518747 Free PMC article.
-
Loss of the Drosophila m-AAA mitochondrial protease paraplegin results in mitochondrial dysfunction, shortened lifespan, and neuronal and muscular degeneration.Cell Death Dis. 2018 Feb 21;9(3):304. doi: 10.1038/s41419-018-0365-8. Cell Death Dis. 2018. PMID: 29467464 Free PMC article.
-
Inactivation of the mitochondrial protease Afg3l2 results in severely diminished respiratory chain activity and widespread defects in mitochondrial gene expression.PLoS Genet. 2020 Oct 19;16(10):e1009118. doi: 10.1371/journal.pgen.1009118. eCollection 2020 Oct. PLoS Genet. 2020. PMID: 33075064 Free PMC article.
-
Lon in maintaining mitochondrial and endoplasmic reticulum homeostasis.Arch Toxicol. 2018 Jun;92(6):1913-1923. doi: 10.1007/s00204-018-2210-3. Epub 2018 May 2. Arch Toxicol. 2018. PMID: 29721585 Review.
-
The peroxisomal Lon protease LonP2 in aging and disease: functions and comparisons with mitochondrial Lon protease LonP1.Biol Rev Camb Philos Soc. 2017 May;92(2):739-753. doi: 10.1111/brv.12253. Epub 2016 Feb 8. Biol Rev Camb Philos Soc. 2017. PMID: 26852705 Free PMC article. Review.
Cited by
-
Evidence for mitochondrial Lonp1 expression in the nucleus.Sci Rep. 2022 Jun 27;12(1):10877. doi: 10.1038/s41598-022-14860-0. Sci Rep. 2022. PMID: 35760833 Free PMC article.
-
Xanthohumol-Induced Rat Glioma C6 Cells Death by Triggering Mitochondrial Stress.Int J Mol Sci. 2021 Apr 26;22(9):4506. doi: 10.3390/ijms22094506. Int J Mol Sci. 2021. PMID: 33925918 Free PMC article.
-
Inactivation of Lon protease reveals a link between mitochondrial unfolded protein stress and mitochondrial translation inhibition.Cell Death Dis. 2018 Dec 5;9(12):1168. doi: 10.1038/s41419-018-1213-6. Cell Death Dis. 2018. PMID: 30518747 Free PMC article.
-
IDH2-mediated regulation of the biogenesis of the oxidative phosphorylation system.Sci Adv. 2022 May 13;8(19):eabl8716. doi: 10.1126/sciadv.abl8716. Epub 2022 May 11. Sci Adv. 2022. PMID: 35544578 Free PMC article.
-
The Role of Lonp1 on Mitochondrial Functions during Cardiovascular and Muscular Diseases.Antioxidants (Basel). 2023 Feb 28;12(3):598. doi: 10.3390/antiox12030598. Antioxidants (Basel). 2023. PMID: 36978846 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
