

The extreme hyper-reactivity of Cys94 in lysozyme avoids its amorphous aggregation
- PMID: 30375487
- PMCID: PMC6207692
- DOI: 10.1038/s41598-018-34439-y
The extreme hyper-reactivity of Cys94 in lysozyme avoids its amorphous aggregation
Abstract
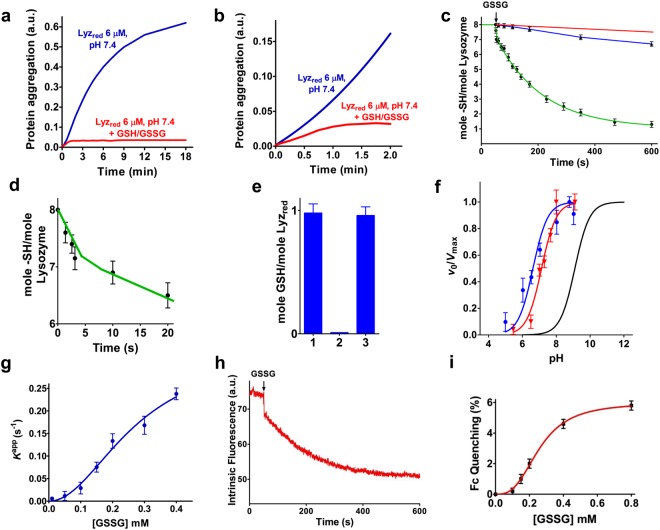
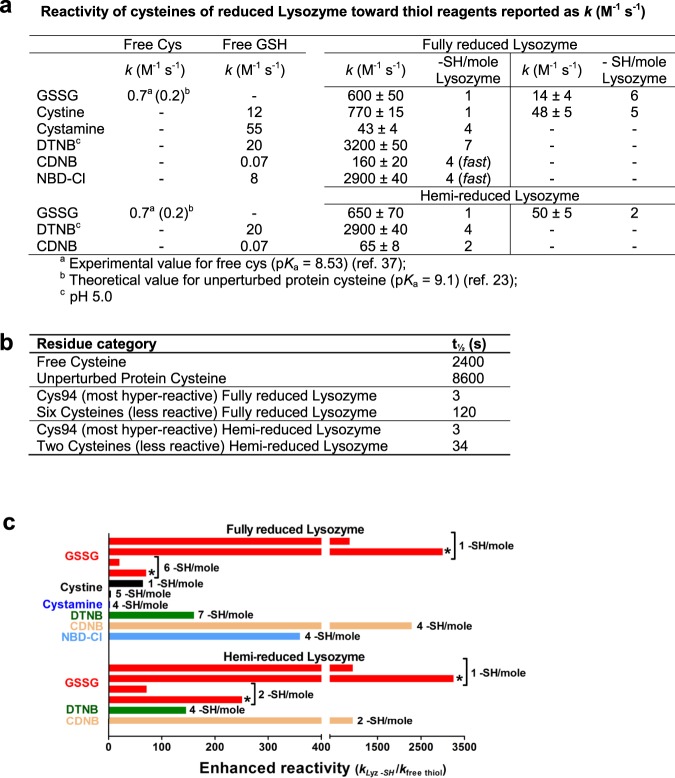
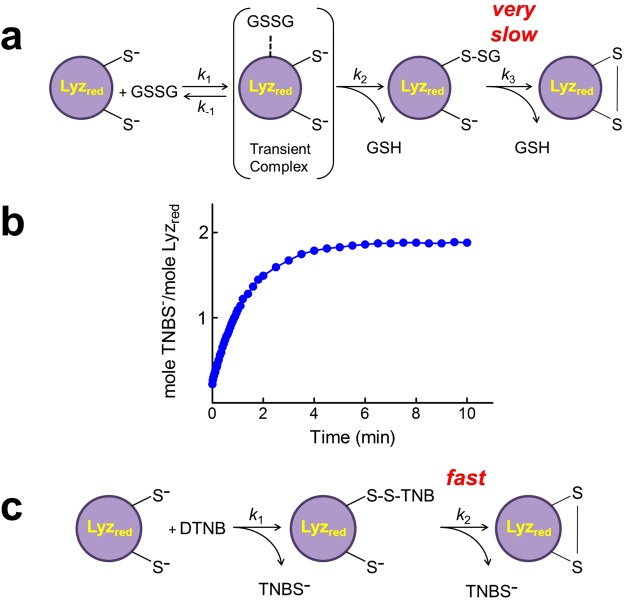
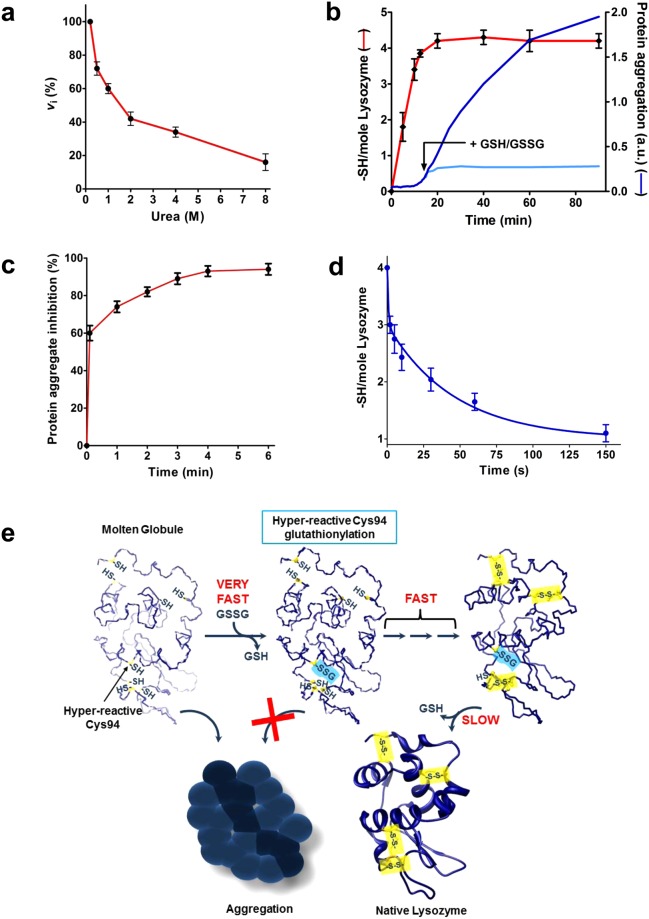
Many proteins provided with disulfide bridges in the native state undergo amorphous irreversible aggregation when these bonds are not formed. Here we show that egg lysozyme displays a clever strategy to prevent this deleterious aggregation during the nascent phase when disulfides are still absent. In fact, when the reduced protein assembles into a molten globule state, its cysteines acquire strong hyper-reactivity towards natural disulfides. The most reactive residue, Cys94, reacts with oxidized glutathione (GSSG) 3000 times faster than an unperturbed protein cysteine. A low pKa of its sulfhydryl group (6.6/7.1) and a productive complex with GSSG (KD = 0.3 mM), causes a fast glutathionylation of this residue (t1/2 = 3 s) and a complete inhibition of the protein aggregation. Other six cysteines display 70 times higher reactivity toward GSSG. The discovery of extreme hyper-reactivity in cysteines only devoted to structural roles opens new research fields for Alzheimer's and Parkinson diseases.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
