

Acetaldehyde dehydrogenase 2 interactions with LDLR and AMPK regulate foam cell formation
- PMID: 30375985
- PMCID: PMC6307968
- DOI: 10.1172/JCI122064
Acetaldehyde dehydrogenase 2 interactions with LDLR and AMPK regulate foam cell formation
Abstract
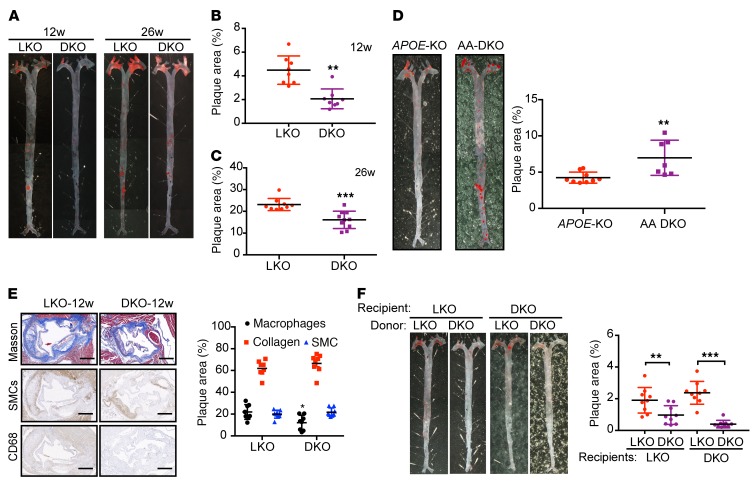
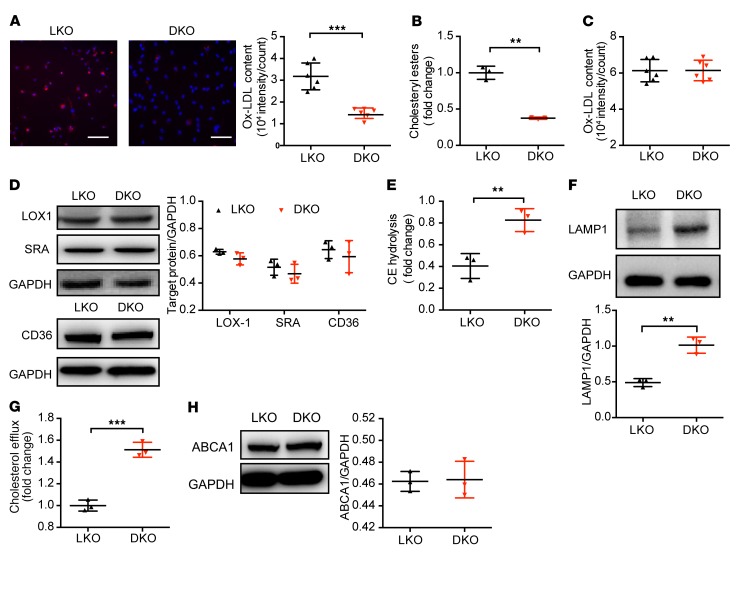
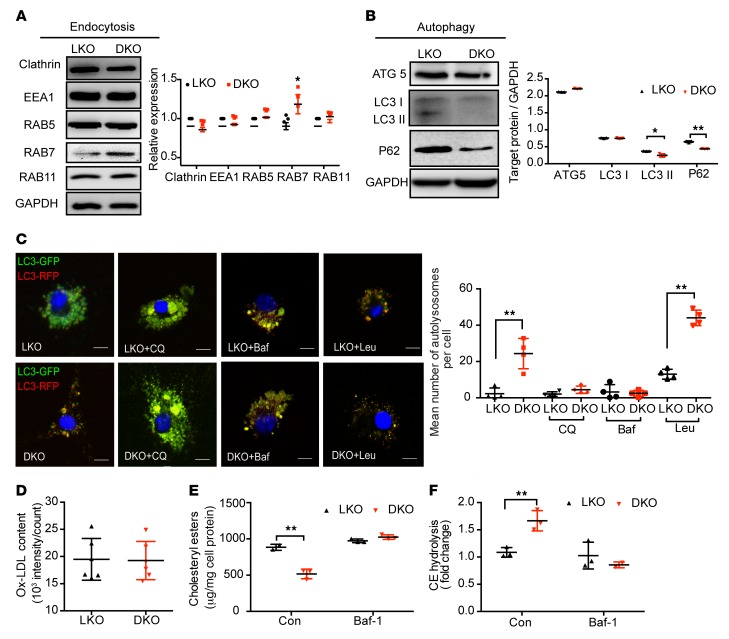
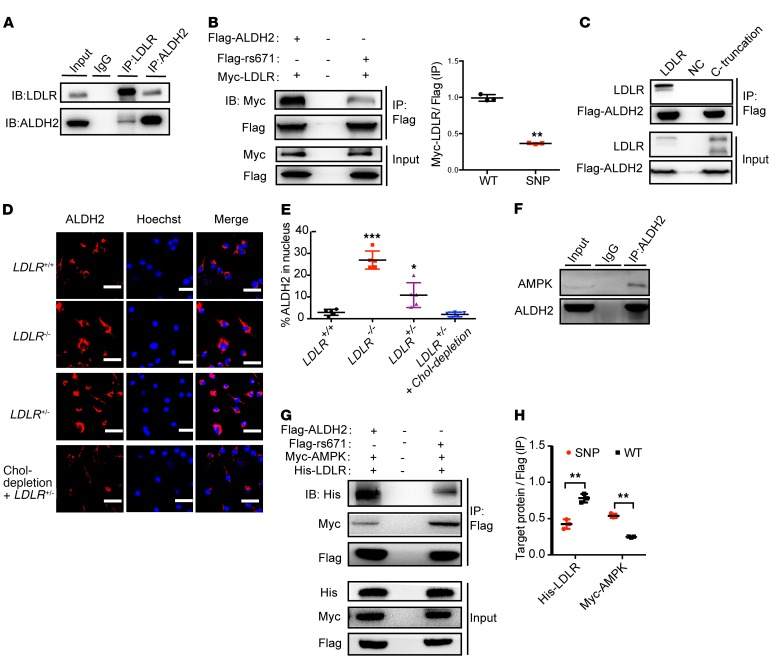
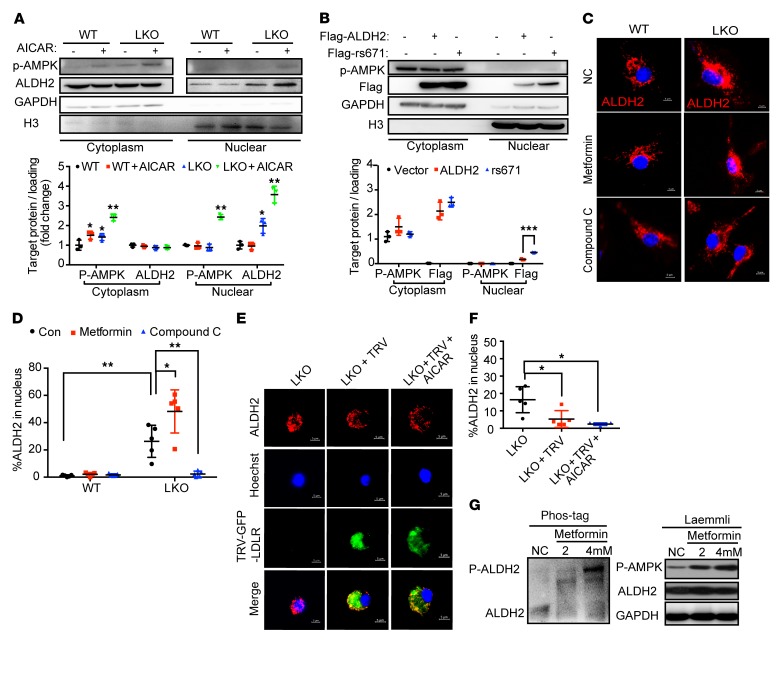
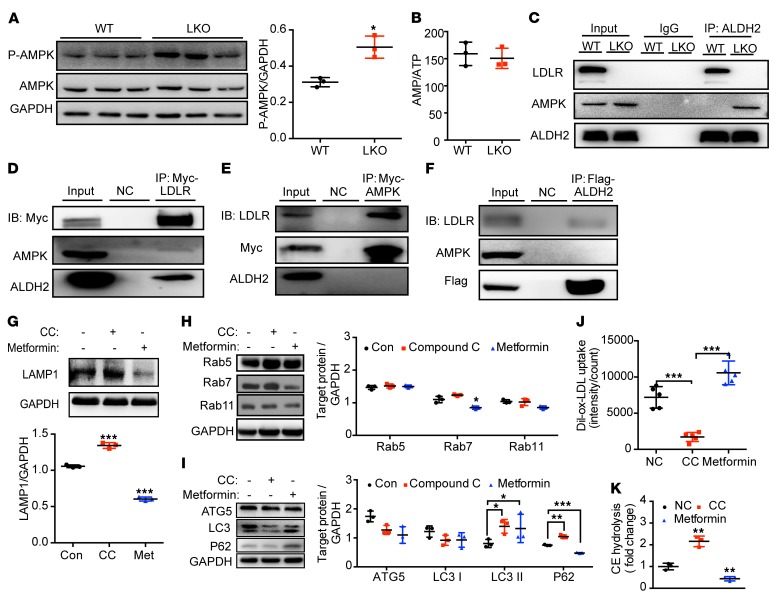
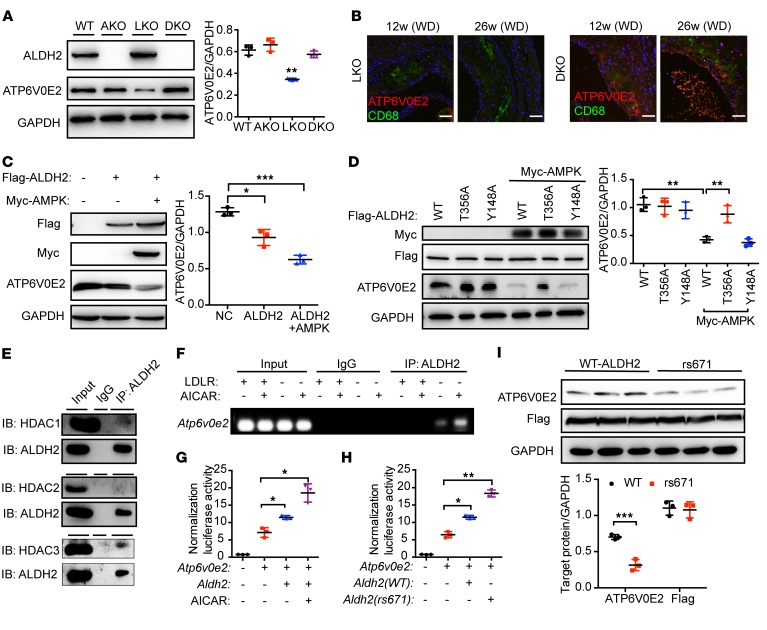
Acetaldehyde dehydrogenase 2 (ALDH2) is a mitochondrial enzyme detoxifying acetaldehyde and endogenous lipid aldehydes; previous studies suggest a protective role of ALDH2 against cardiovascular disease (CVD). Around 40% of East Asians carrying the single nucleotide polymorphism (SNP) ALDH2 rs671 have an increased incidence of CVD. However, the role of ALDH2 in CVD beyond alcohol consumption remains poorly defined. Here we report that ALDH2/LDLR double knockout (DKO) mice have decreased atherosclerosis compared with LDLR-KO mice, whereas ALDH2/APOE-DKO mice have increased atherosclerosis, suggesting an unexpected interaction of ALDH2 with LDLR. Further studies demonstrate that in the absence of LDLR, AMPK phosphorylates ALDH2 at threonine 356 and enables its nuclear translocation. Nuclear ALDH2 interacts with HDAC3 and represses transcription of a lysosomal proton pump protein ATP6V0E2, critical for maintaining lysosomal function, autophagy, and degradation of oxidized low-density lipid protein. Interestingly, an interaction of cytosolic LDLR C-terminus with AMPK blocks ALDH2 phosphorylation and subsequent nuclear translocation, whereas ALDH2 rs671 mutant in human macrophages attenuates this interaction, which releases ALDH2 to the nucleus to suppress ATP6V0E2 expression, resulting in increased foam cells due to impaired lysosomal function. Our studies reveal a novel role of ALDH2 and LDLR in atherosclerosis and provide a molecular mechanism by which ALDH2 rs671 SNP increases CVD.
Keywords: Atherosclerosis; Cardiovascular disease; Cell Biology; Macrophages; Metabolism.
Conflict of interest statement
Figures


Comment in
-
Not just correlative: a new pathway defines how an ALDH2 SNP contributes to atherosclerosis.J Clin Invest. 2019 Jan 2;129(1):63-65. doi: 10.1172/JCI125433. Epub 2018 Dec 3. J Clin Invest. 2019. PMID: 30507608 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
