

Identifying deer antler uhrf1 proliferation and s100a10 mineralization genes using comparative RNA-seq
- PMID: 30376879
- PMCID: PMC6208050
- DOI: 10.1186/s13287-018-1027-6
Identifying deer antler uhrf1 proliferation and s100a10 mineralization genes using comparative RNA-seq
Abstract
Background: Deer antlers are bony structures that re-grow at very high rates, making them an attractive model for studying rapid bone regeneration.
Methods: To identify the genes that are involved in this fast pace of bone growth, an in vitro RNA-seq model that paralleled the sharp differences in bone growth between deer antlers and humans was established. Subsequently, RNA-seq (> 60 million reads per library) was used to compare transcriptomic profiles. Uniquely expressed deer antler proliferation as well as mineralization genes were identified via a combination of differential gene expression and subtraction analysis. Thereafter, the physiological relevance as well as contributions of these identified genes were determined by immunofluorescence, gene overexpression, and gene knockdown studies.
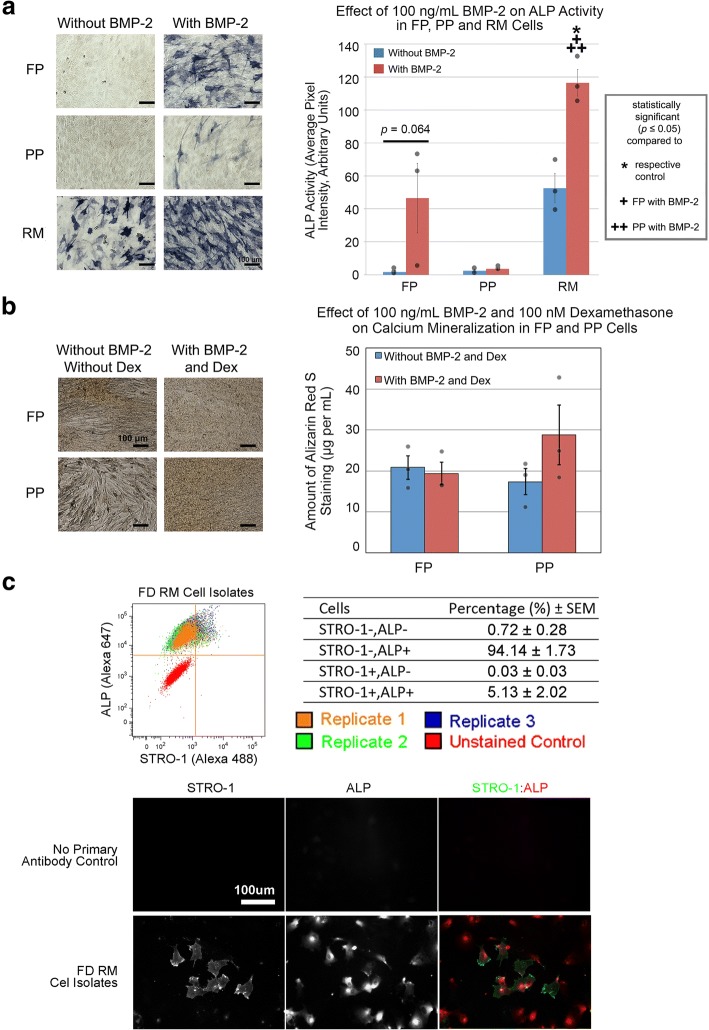
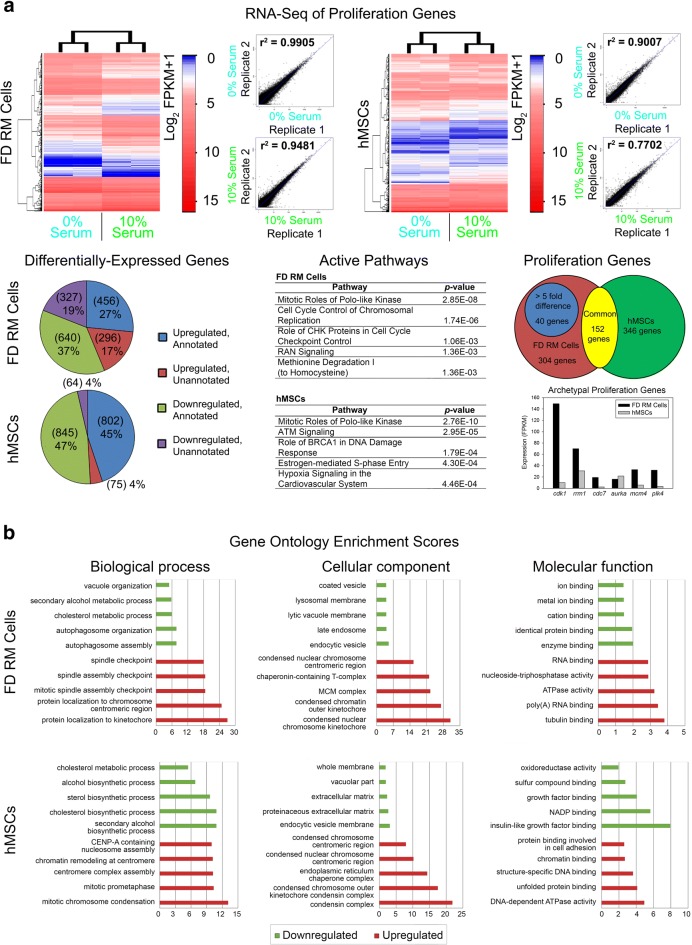
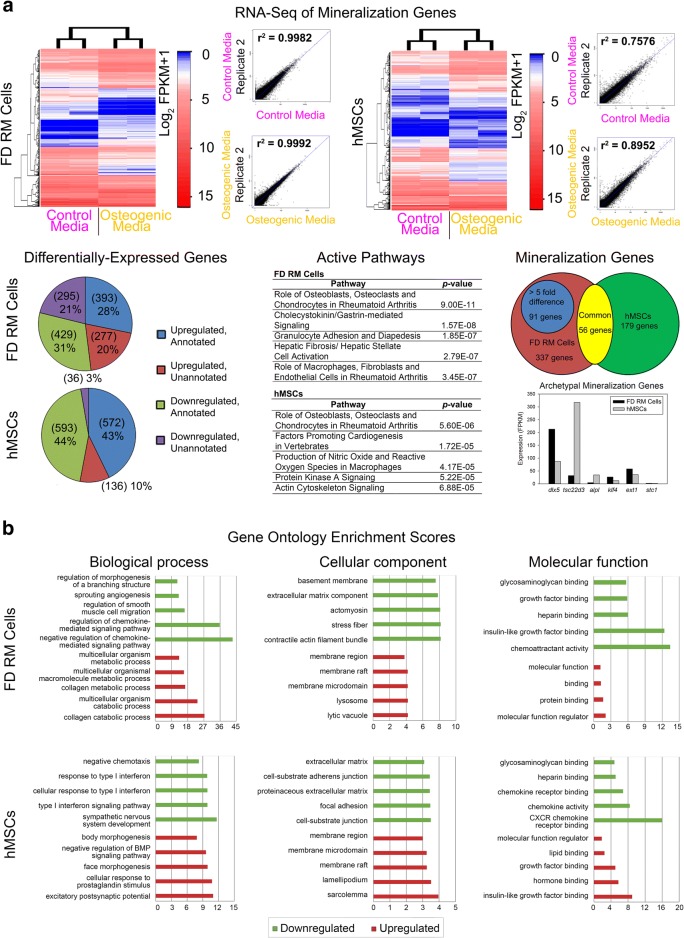
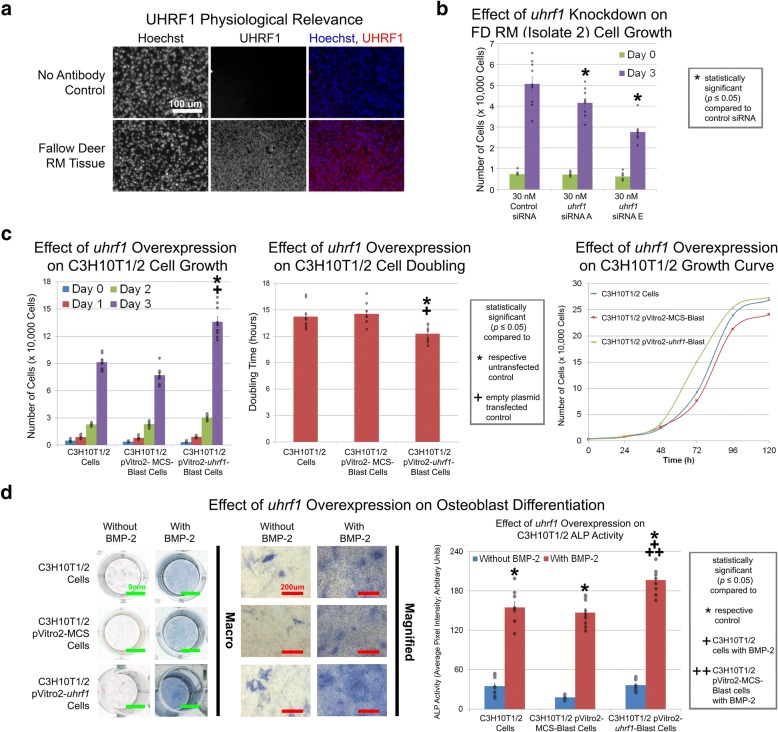
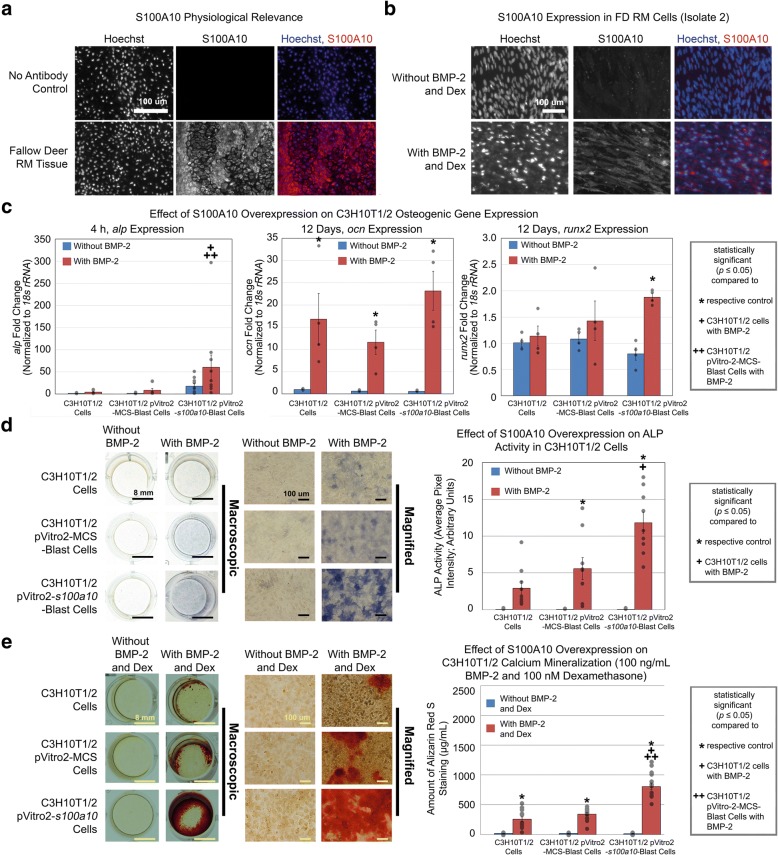
Results: Cell characterization studies showed that in vitro-cultured deer antler-derived reserve mesenchyme (RM) cells exhibited high osteogenic capabilities and cell surface markers similar to in vivo counterparts. Under identical culture conditions, deer antler RM cells proliferated faster (8.6-11.7-fold increase in cell numbers) and exhibited increased osteogenic differentiation (17.4-fold increase in calcium mineralization) compared to human mesenchymal stem cells (hMSCs), paralleling in vivo conditions. Comparative RNA-seq identified 40 and 91 previously unknown and uniquely expressed fallow deer (FD) proliferation and mineralization genes, respectively, including uhrf1 and s100a10. Immunofluorescence studies showed that uhrf1 and s100a10 were expressed in regenerating deer antlers while gene overexpression and gene knockdown studies demonstrated the proliferation contributions of uhrf1 and mineralization capabilities of s100a10.
Conclusion: Using a simple, in vitro comparative RNA-seq approach, novel genes pertinent to fast bony antler regeneration were identified and their proliferative/osteogenic function was verified via gene overexpression, knockdown, and immunostaining. This combinatorial approach may be applicable to discover unique gene contributions between any two organisms for a given phenomenon-of-interest.
Keywords: Bone regeneration; Comparative RNA-seq; Deer antler; s100a10; uhrf1.
Conflict of interest statement
Ethics approval
Ethics approval for the harvesting of deer antlers was obtained from Stanford University’s Administrative Panel on Laboratory Animal Care (APLAC 28057).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures

References
-
- Gómez JA, Ceacero F, Landete-Castillejos T, Gaspar-López E, García AJ, Gallego L. Factors affecting antler investment in Iberian red deer. Anim Prod Sci. 2012;52:867–873. doi: 10.1071/AN11316. - DOI
-
- Huxley JS. The relative size of antlers in deer. Proc Zool Soc London. 1931;101:819–864. doi: 10.1111/j.1096-3642.1931.tb01047.x. - DOI
-
- Goss RJ. Deer antlers. Regeneration, function and evolution. New York: Academic Press; 1983.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
