

CLASP stabilization of plus ends created by severing promotes microtubule creation and reorientation
- PMID: 30377221
- PMCID: PMC6314540
- DOI: 10.1083/jcb.201805047
CLASP stabilization of plus ends created by severing promotes microtubule creation and reorientation
Abstract
Central to the building and reorganizing cytoskeletal arrays is creation of new polymers. Although nucleation has been the major focus of study for microtubule generation, severing has been proposed as an alternative mechanism to create new polymers, a mechanism recently shown to drive the reorientation of cortical arrays of higher plants in response to blue light perception. Severing produces new plus ends behind the stabilizing GTP-cap. An important and unanswered question is how these ends are stabilized in vivo to promote net microtubule generation. Here we identify the conserved protein CLASP as a potent stabilizer of new plus ends created by katanin severing in plant cells. Clasp mutants are defective in cortical array reorientation. In these mutants, both rescue of shrinking plus ends and the stabilization of plus ends immediately after severing are reduced. Computational modeling reveals that it is the specific stabilization of severed ends that best explains CLASP's function in promoting microtubule amplification by severing and array reorientation.
© 2018 Lindeboom et al.
Figures
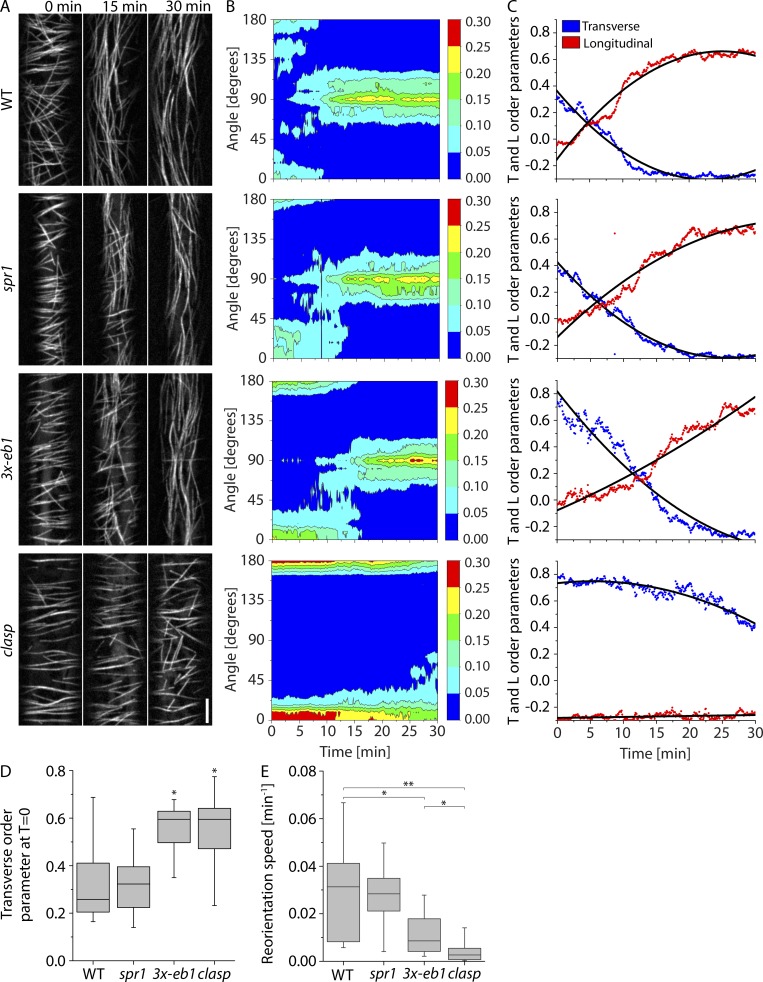
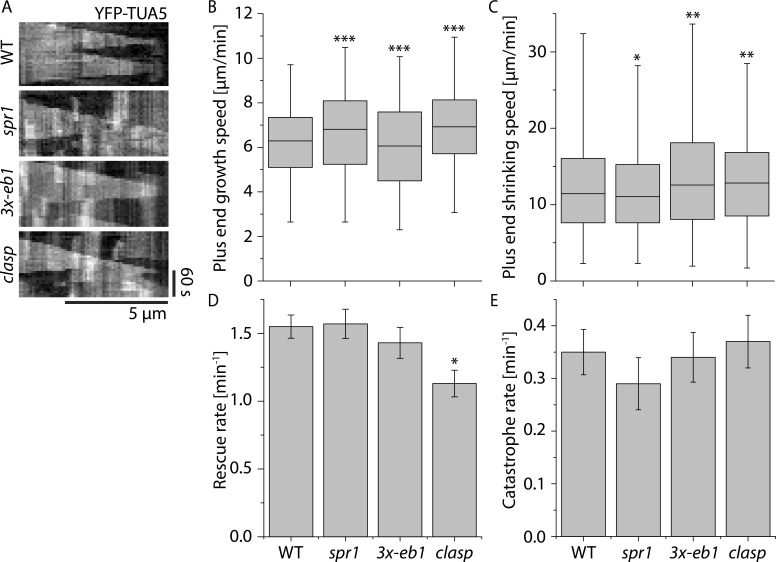
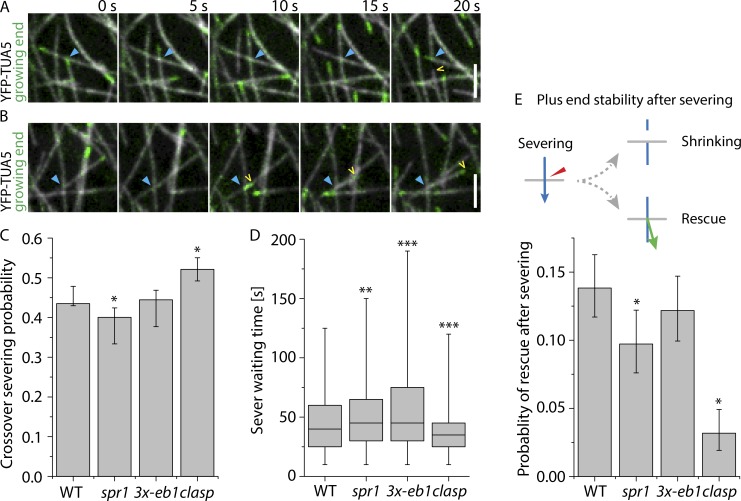
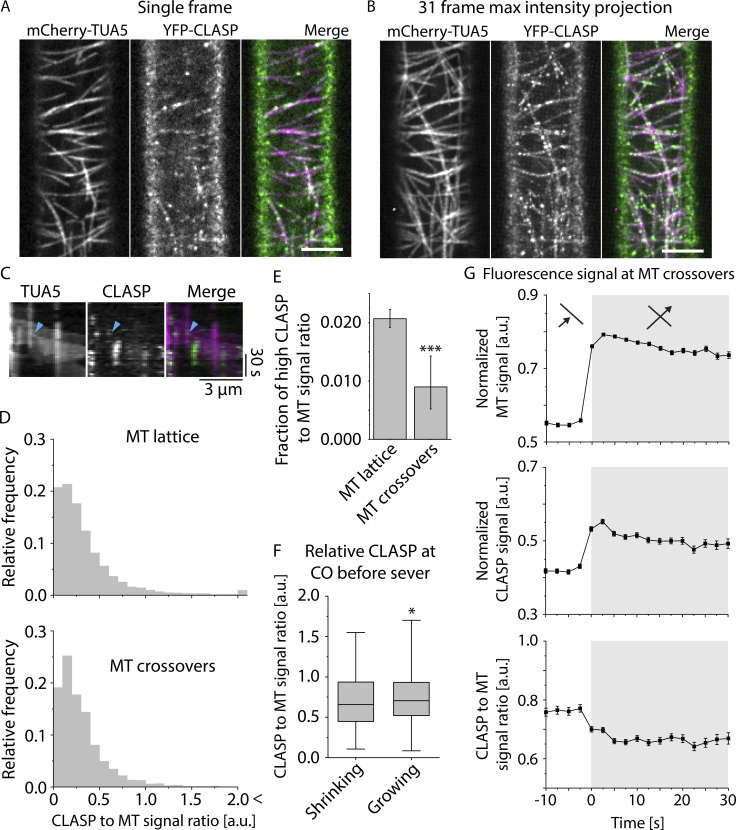
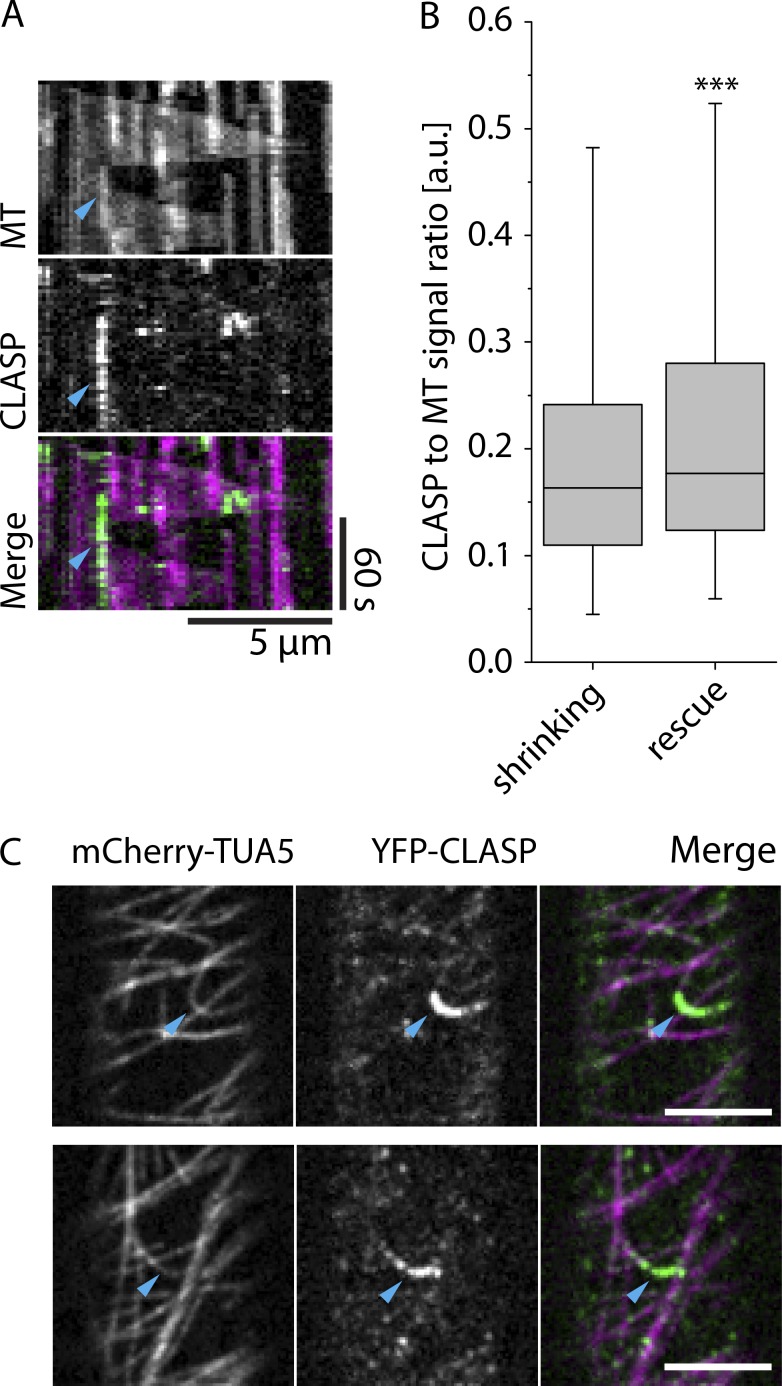
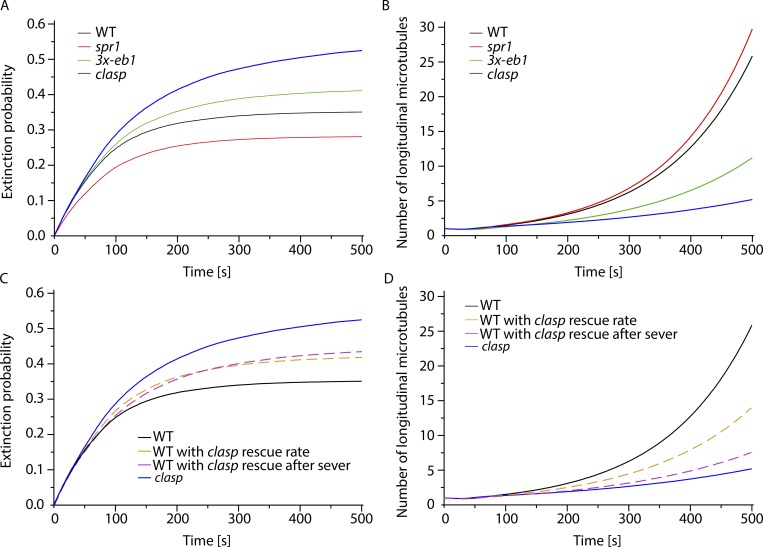
Comment in
-
Microtubule reorientation in the blue spotlight: Cutting and CLASPing at dynamic hot spots.J Cell Biol. 2019 Jan 7;218(1):8-9. doi: 10.1083/jcb.201812063. Epub 2018 Dec 20. J Cell Biol. 2019. PMID: 30573524 Free PMC article.
References
-
- Abramowitz M., and Stegun I.A.. 1965. Handbook of mathematical functions with formulas, graphs, and mathematical tables. Dover Publications, New York. 1 pp.
-
- Akhmanova A., Hoogenraad C.C., Drabek K., Stepanova T., Dortland B., Verkerk T., Vermeulen W., Burgering B.M., De Zeeuw C.I., Grosveld F., and Galjart N.. 2001. Clasps are CLIP-115 and -170 associating proteins involved in the regional regulation of microtubule dynamics in motile fibroblasts. Cell. 104:923–935. 10.1016/S0092-8674(01)00288-4 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
