

Evolution and development of the fish jaw skeleton
- PMID: 30378758
- PMCID: PMC8299565
- DOI: 10.1002/wdev.337
Evolution and development of the fish jaw skeleton
Abstract
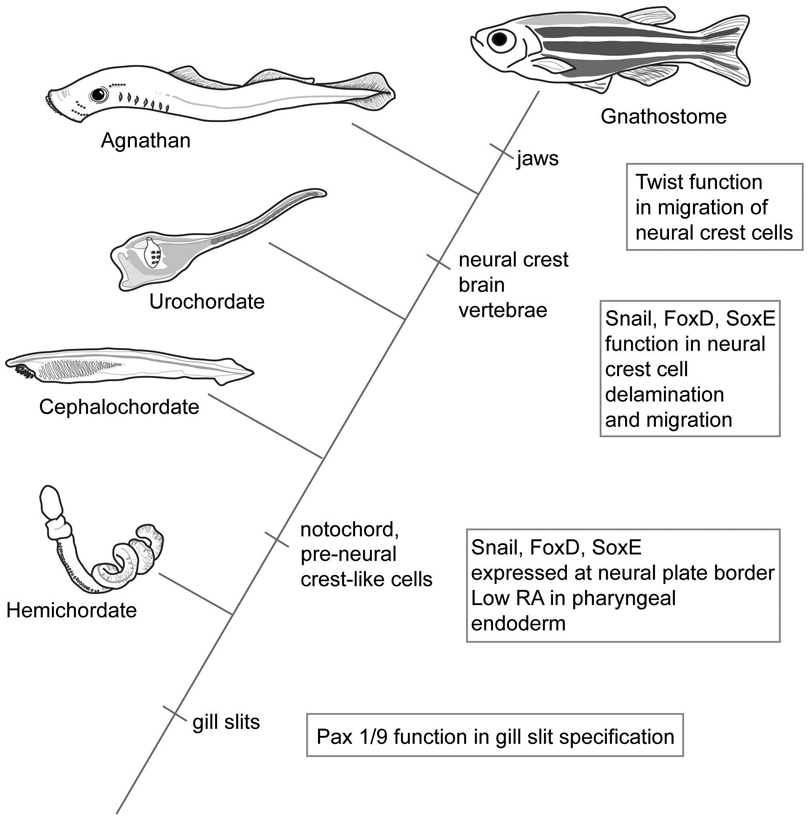
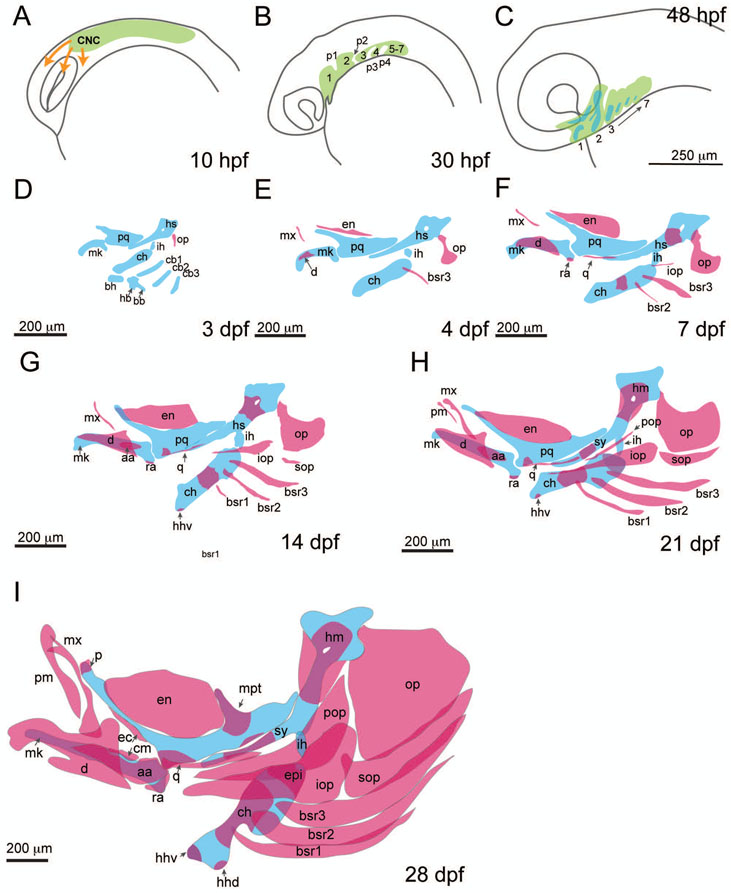
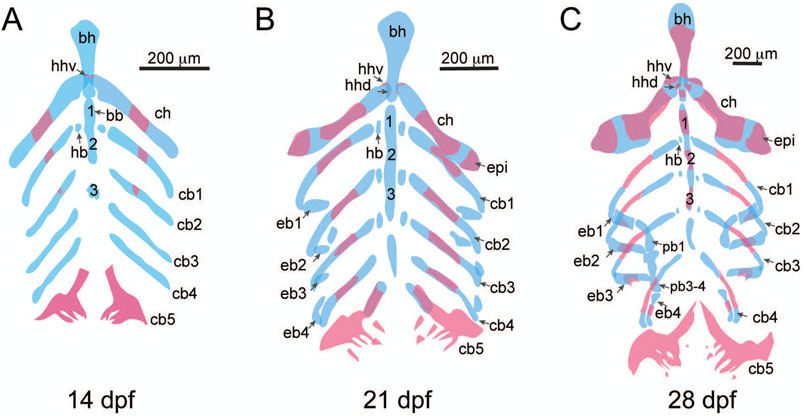
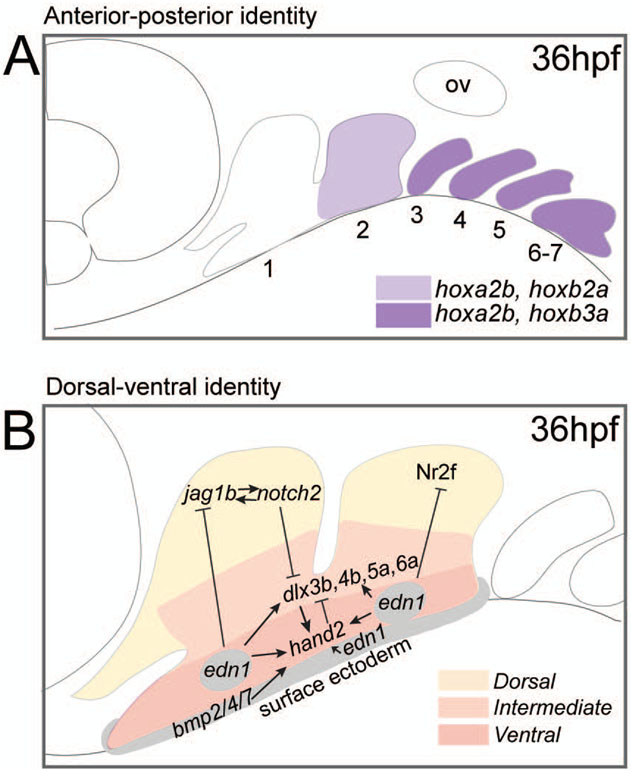
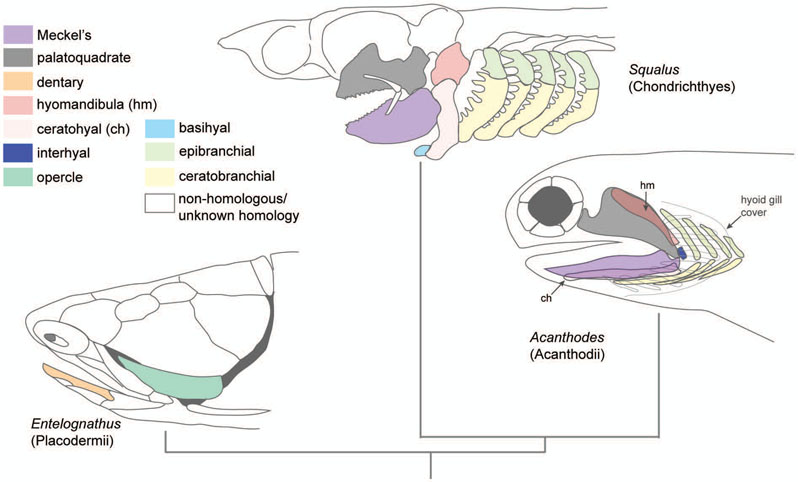
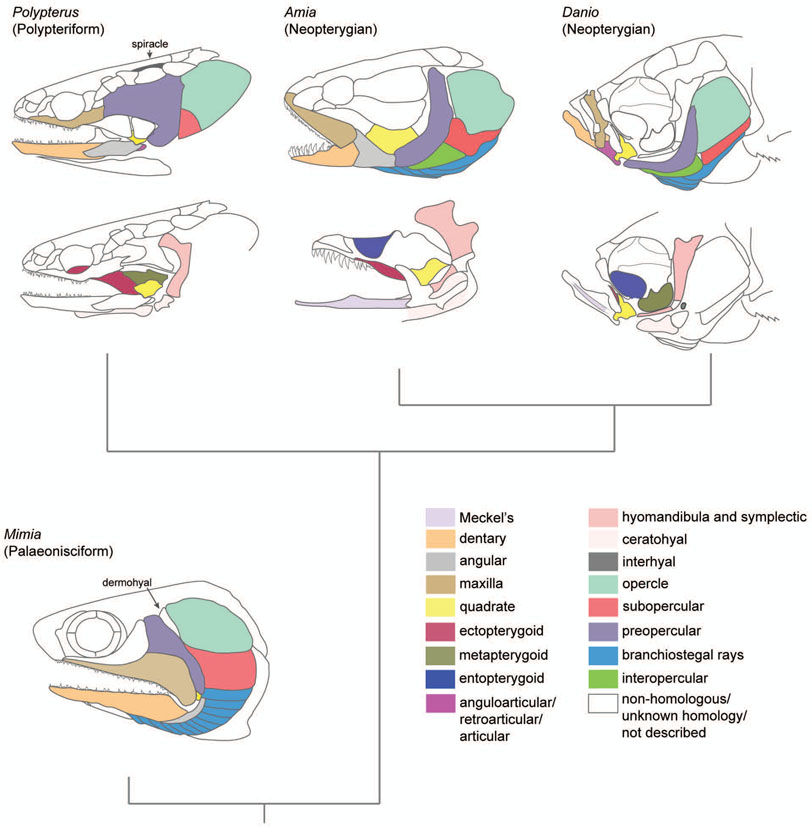
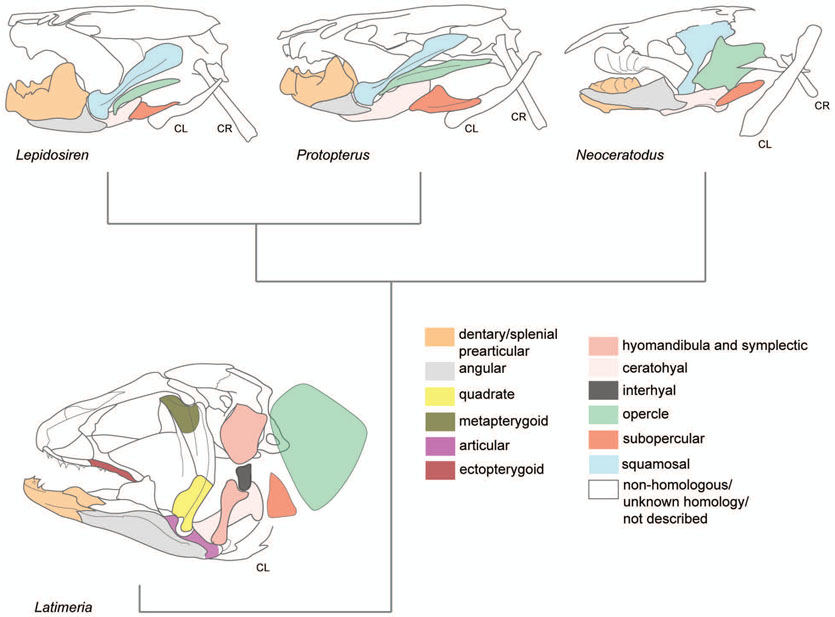
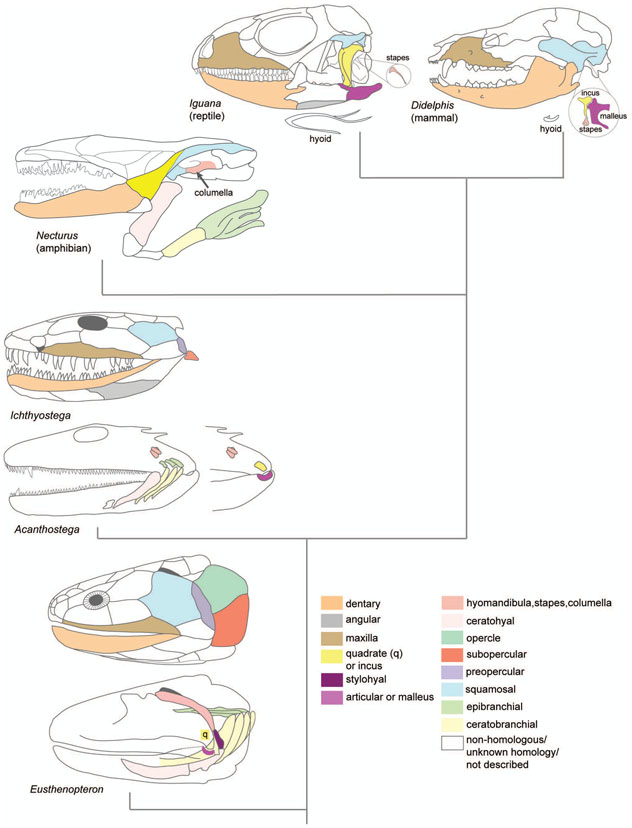
The evolution of the jaw represents a key innovation in driving the diversification of vertebrate body plans and behavior. The pharyngeal apparatus originated as gill bars separated by slits in chordate ancestors to vertebrates. Later, with the acquisition of neural crest, pharyngeal arches gave rise to branchial basket cartilages in jawless vertebrates (agnathans), and later bone and cartilage of the jaw, jaw support, and gills of jawed vertebrates (gnathostomes). Major events in the evolution of jaw structure from agnathans to gnathostomes include axial regionalization of pharyngeal elements and formation of a jaw joint. Hox genes specify the anterior-posterior identity of arches, and edn1, dlx, hand2, Jag1b-Notch2 signaling, and Nr2f factors specify dorsal-ventral identity. The formation of a jaw joint, an important step in the transition from an un-jointed pharynx in agnathans to a hinged jaw in gnathostomes involves interaction between nkx3.2, hand2, and barx1 factors. Major events in jaw patterning between fishes and reptiles include changes to elements of the second pharyngeal arch, including a loss of opercular and branchiostegal ray bones and transformation of the hyomandibula into the stapes. Further changes occurred between reptiles and mammals, including the transformation of the articular and quadrate elements of the jaw joint into the malleus and incus of the middle ear. Fossils of transitional jaw phenotypes can be analyzed from a developmental perspective, and there exists potential to use genetic manipulation techniques in extant taxa to test hypotheses about the evolution of jaw patterning in ancient vertebrates. This article is categorized under: Comparative Development and Evolution > Evolutionary Novelties Early Embryonic Development > Development to the Basic Body Plan Comparative Development and Evolution > Body Plan Evolution.
Keywords: jaw; morphogenesis; neural crest; skeleton; vertebrates.
© 2018 Wiley Periodicals, Inc.
Conflict of interest statement
No conflicts of interest
Figures
Similar articles
-
Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint.Development. 2003 Apr;130(7):1353-65. doi: 10.1242/dev.00339. Development. 2003. PMID: 12588851
-
Bapx1 regulates patterning in the middle ear: altered regulatory role in the transition from the proximal jaw during vertebrate evolution.Development. 2004 Mar;131(6):1235-45. doi: 10.1242/dev.01017. Epub 2004 Feb 18. Development. 2004. PMID: 14973294
-
Fishing for jaws in early vertebrate evolution: a new hypothesis of mandibular confinement.Biol Rev Camb Philos Soc. 2016 Aug;91(3):611-57. doi: 10.1111/brv.12187. Epub 2015 Apr 21. Biol Rev Camb Philos Soc. 2016. PMID: 25899041 Review.
-
Evidence for the prepattern/cooption model of vertebrate jaw evolution.Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17262-7. doi: 10.1073/pnas.1009304107. Epub 2010 Sep 20. Proc Natl Acad Sci U S A. 2010. PMID: 20855630 Free PMC article.
-
Neural crest patterning and the evolution of the jaw.J Anat. 2001 Jul-Aug;199(Pt 1-2):105-20. doi: 10.1046/j.1469-7580.2001.19910105.x. J Anat. 2001. PMID: 11523812 Free PMC article. Review.
Cited by
-
Phenotypic covariation predicts diversification in an adaptive radiation of pupfishes.Ecol Evol. 2024 Aug 7;14(8):e11642. doi: 10.1002/ece3.11642. eCollection 2024 Aug. Ecol Evol. 2024. PMID: 39114171 Free PMC article.
-
Identification of signalling pathways involved in gill regeneration in zebrafish.J Exp Biol. 2024 Jan 15;227(2):jeb246290. doi: 10.1242/jeb.246290. Epub 2024 Jan 26. J Exp Biol. 2024. PMID: 38099598 Free PMC article.
-
Deficiency in the endocytic adaptor proteins PHETA1/2 impairs renal and craniofacial development.Dis Model Mech. 2020 May 26;13(5):dmm041913. doi: 10.1242/dmm.041913. Dis Model Mech. 2020. PMID: 32152089 Free PMC article.
-
Nkx2.7 is a conserved regulator of craniofacial development.Nat Commun. 2025 Apr 23;16(1):3802. doi: 10.1038/s41467-025-58821-3. Nat Commun. 2025. PMID: 40268889 Free PMC article.
-
Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome.Cells. 2022 Apr 8;11(8):1260. doi: 10.3390/cells11081260. Cells. 2022. PMID: 35455940 Free PMC article. Review.
References
-
- Amores A, Force A, Yan YL, Joly L, Amemiya C, Fritz A, … Postlethwait JH (1998). Zebrafish hox clusters and vertebrate genome evolution. Science (New York, N.Y.), 282(5394), 1711–1714. - PubMed
Further Reading
-
- Clack JA (2002). Gaining Ground: The Origin and Evolution of Tetrapods. Indiana University Press.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
