

CD16+ monocytes give rise to CD103+RALDH2+TCF4+ dendritic cells with unique transcriptional and immunological features
- PMID: 30381402
- PMCID: PMC6234376
- DOI: 10.1182/bloodadvances.2018020123
CD16+ monocytes give rise to CD103+RALDH2+TCF4+ dendritic cells with unique transcriptional and immunological features
Abstract
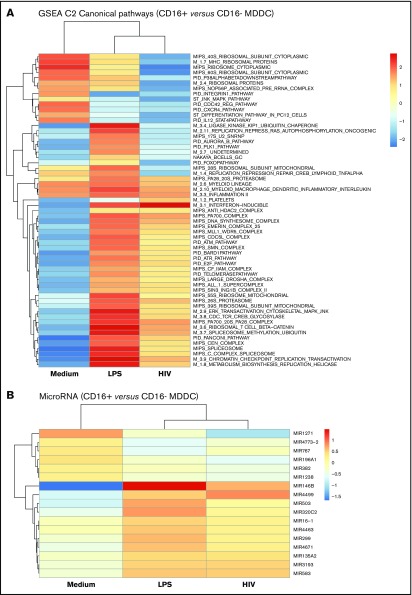
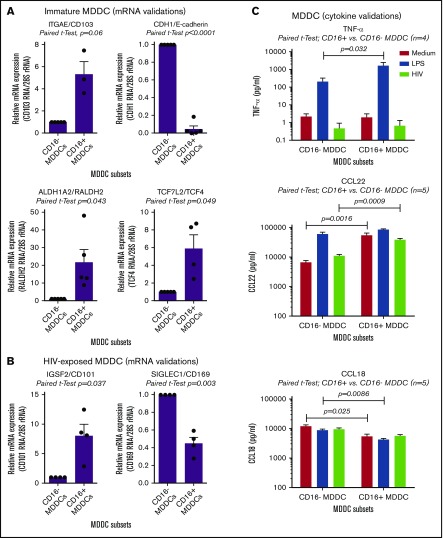
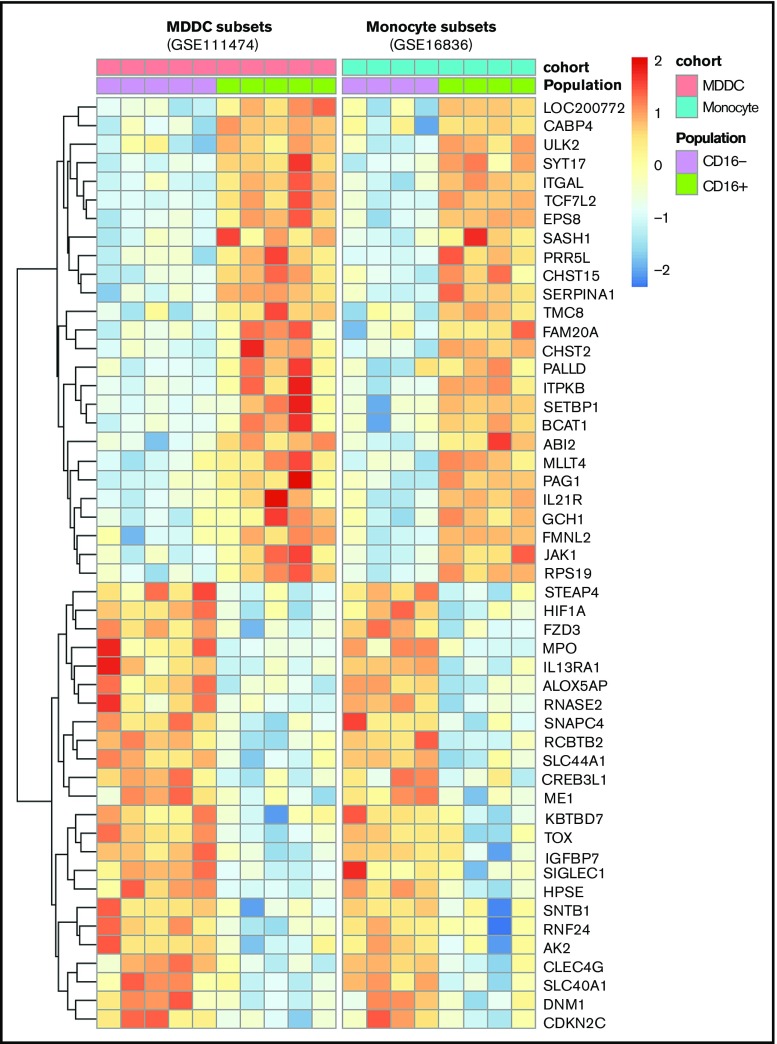
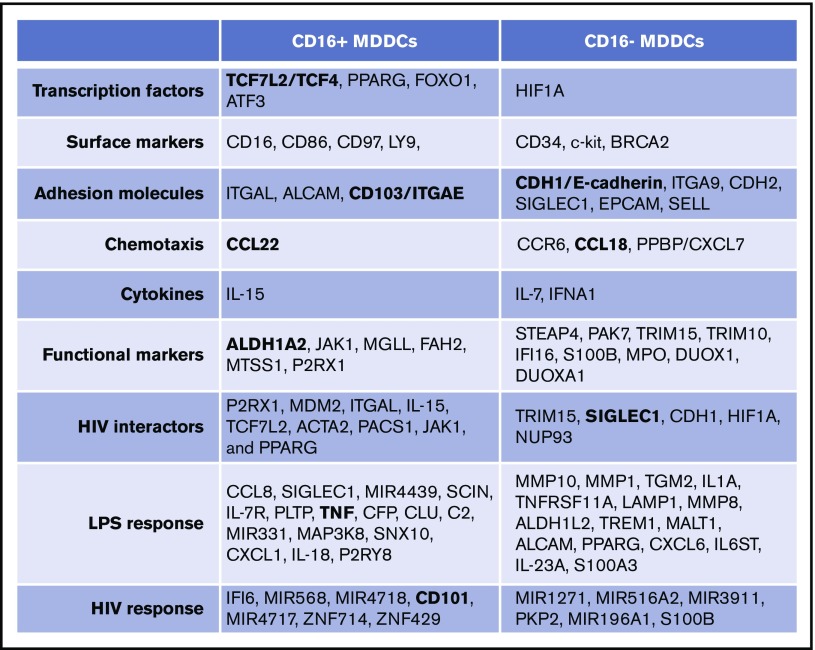
Classical CD16- vs intermediate/nonclassical CD16+ monocytes differ in their homing potential and biological functions, but whether they differentiate into dendritic cells (DCs) with distinct contributions to immunity against bacterial/viral pathogens remains poorly investigated. Here, we employed a systems biology approach to identify clinically relevant differences between CD16+ and CD16- monocyte-derived DCs (MDDCs). Although both CD16+ and CD16- MDDCs acquire classical immature/mature DC markers in vitro, genome-wide transcriptional profiling revealed unique molecular signatures for CD16+ MDDCs, including adhesion molecules (ITGAE/CD103), transcription factors (TCF7L2/TCF4), and enzymes (ALDH1A2/RALDH2), whereas CD16- MDDCs exhibit a CDH1/E-cadherin+ phenotype. Of note, lipopolysaccharides (LPS) upregulated distinct transcripts in CD16+ (eg, CCL8, SIGLEC1, MIR4439, SCIN, interleukin [IL]-7R, PLTP, tumor necrosis factor [TNF]) and CD16- MDDCs (eg, MMP10, MMP1, TGM2, IL-1A, TNFRSF11A, lysosomal-associated membrane protein 1, MMP8). Also, unique sets of HIV-modulated genes were identified in the 2 subsets. Further gene set enrichment analysis identified canonical pathways that pointed to "inflammation" as the major feature of CD16+ MDDCs at immature stage and on LPS/HIV exposure. Finally, functional validations and meta-analysis comparing the transcriptome of monocyte and MDDC subsets revealed that CD16+ vs CD16- monocytes preserved their superior ability to produce TNF-α and CCL22, as well as other sets of transcripts (eg, TCF4), during differentiation into DC. These results provide evidence that monocyte subsets are transcriptionally imprinted/programmed with specific differentiation fates, with intermediate/nonclassical CD16+ monocytes being precursors for pro-inflammatory CD103+RALDH2+TCF4+ DCs that may play key roles in mucosal immunity homeostasis/pathogenesis. Thus, alterations in the CD16+ /CD16- monocyte ratios during pathological conditions may dramatically influence the quality of MDDC-mediated immunity.
© 2018 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures

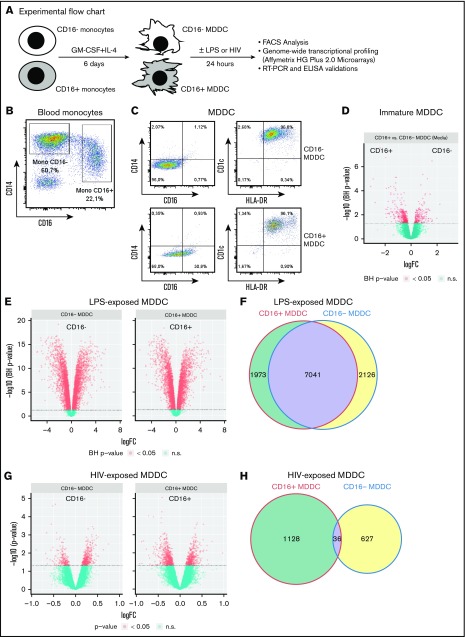
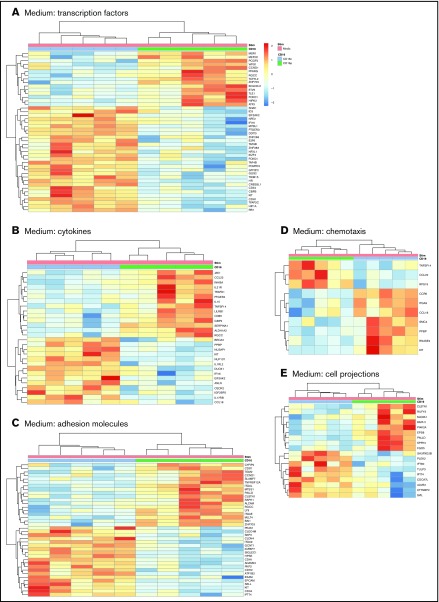
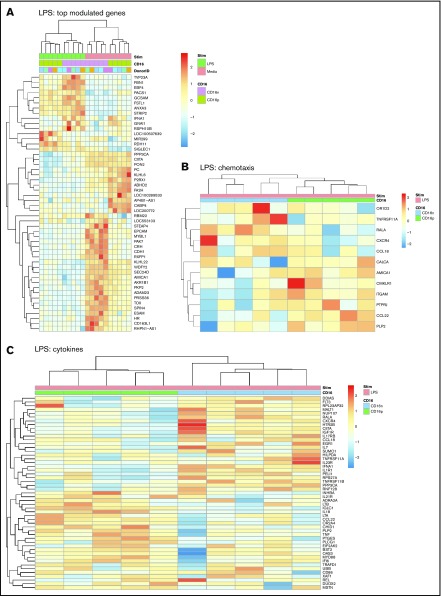
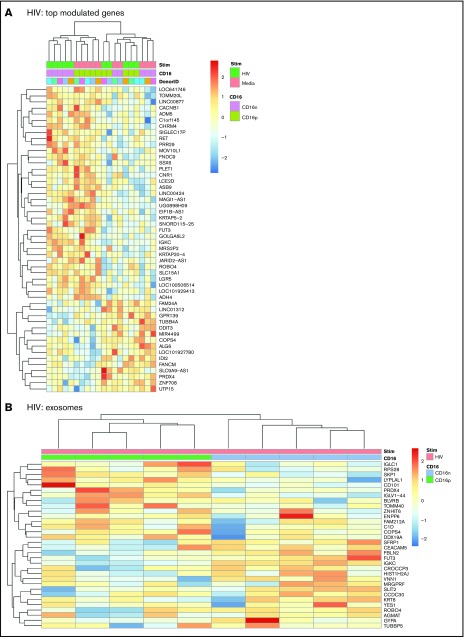
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
