

Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice
- PMID: 30381407
- PMCID: PMC6483075
- DOI: 10.1126/scitranslmed.aah4066
Inflammasome inhibition prevents α-synuclein pathology and dopaminergic neurodegeneration in mice
Abstract
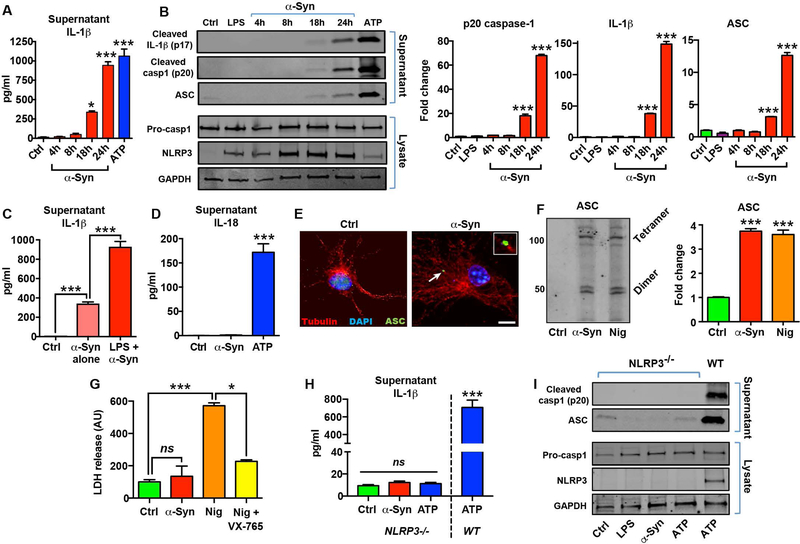
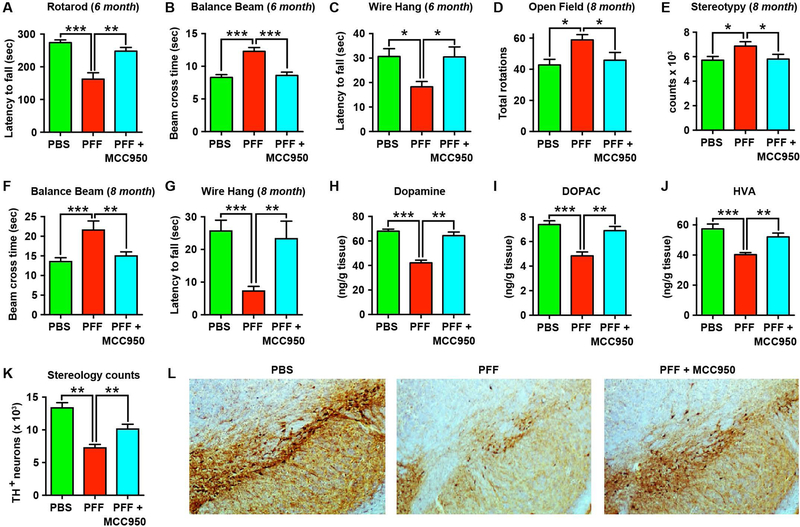
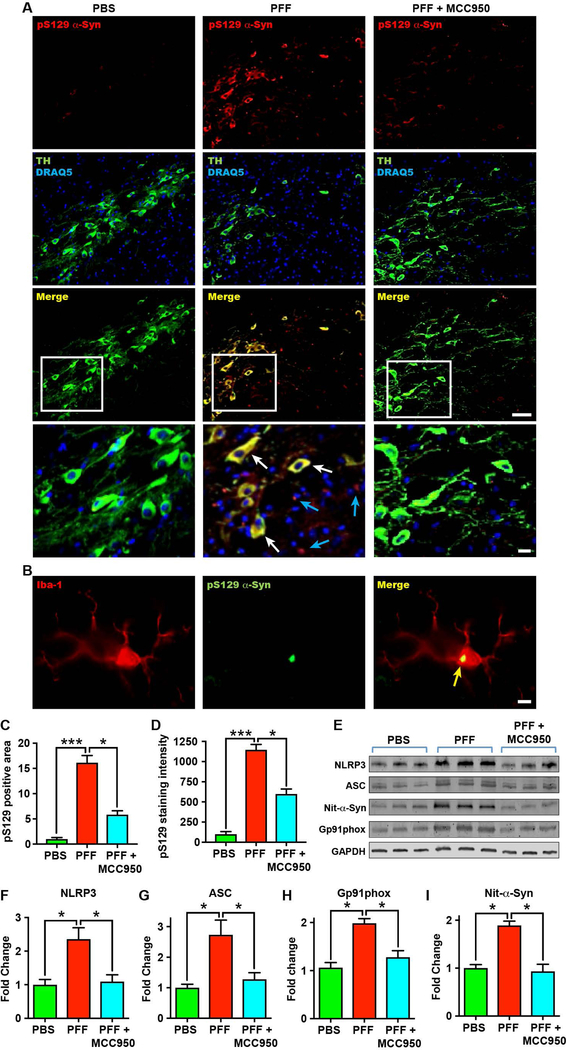
Parkinson's disease (PD) is characterized by a profound loss of dopaminergic neurons in the substantia nigra, accompanied by chronic neuroinflammation, mitochondrial dysfunction, and widespread accumulation of α-synuclein-rich protein aggregates in the form of Lewy bodies. However, the mechanisms linking α-synuclein pathology and dopaminergic neuronal death to chronic microglial neuroinflammation have not been completely elucidated. We show that activation of the microglial NLR family pyrin domain containing 3 (NLRP3) inflammasome is a common pathway triggered by both fibrillar α-synuclein and dopaminergic degeneration in the absence of α-synuclein aggregates. Cleaved caspase-1 and the inflammasome adaptor protein apoptosis-associated speck-like protein containing a C-terminal caspase recruitment domain (ASC) were elevated in the substantia nigra of the brains of patients with PD and in multiple preclinical PD models. NLRP3 activation by fibrillar α-synuclein in mouse microglia resulted in a delayed but robust activation of the NLRP3 inflammasome leading to extracellular interleukin-1β and ASC release in the absence of pyroptosis. Nanomolar doses of a small-molecule NLRP3 inhibitor, MCC950, abolished fibrillar α-synuclein-mediated inflammasome activation in mouse microglial cells and extracellular ASC release. Furthermore, oral administration of MCC950 in multiple rodent PD models inhibited inflammasome activation and effectively mitigated motor deficits, nigrostriatal dopaminergic degeneration, and accumulation of α-synuclein aggregates. These findings suggest that microglial NLRP3 may be a sustained source of neuroinflammation that could drive progressive dopaminergic neuropathology and highlight NLRP3 as a potential target for disease-modifying treatments for PD.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures



References
-
- Poewe W, Seppi K, Tanner CM, Halliday GM, Brundin P, Volkmann J, Schrag AE, Lang AE, Parkinson disease. Nat Rev Dis Primers 3, 17013 (2017). - PubMed
-
- Obeso JA, Rodriguez-Oroz MC, Goetz CG, Marin C, Kordower JH, Rodriguez M, Hirsch EC, Farrer M, Schapira AH, Halliday G, Missing pieces in the Parkinson’s disease puzzle. Nat Med 16, 653–661 (2010). - PubMed
-
- Peelaerts W, Bousset L, Van der Perren A, Moskalyuk A, Pulizzi R, Giugliano M, Van den Haute C, Melki R, Baekelandt V, alpha-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature 522, 340–344 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
