

Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection
- PMID: 30381458
- PMCID: PMC6243247
- DOI: 10.1073/pnas.1809548115
Caspase-8 induces cleavage of gasdermin D to elicit pyroptosis during Yersinia infection
Abstract
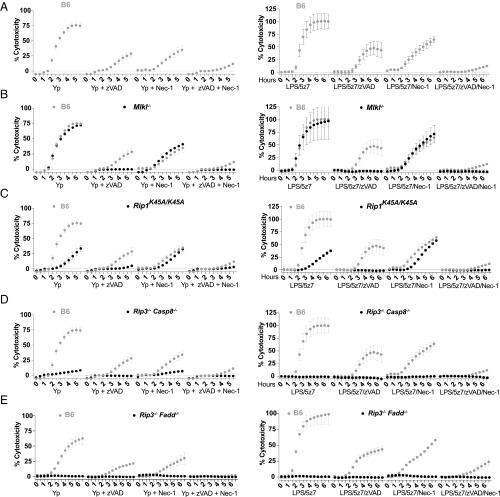
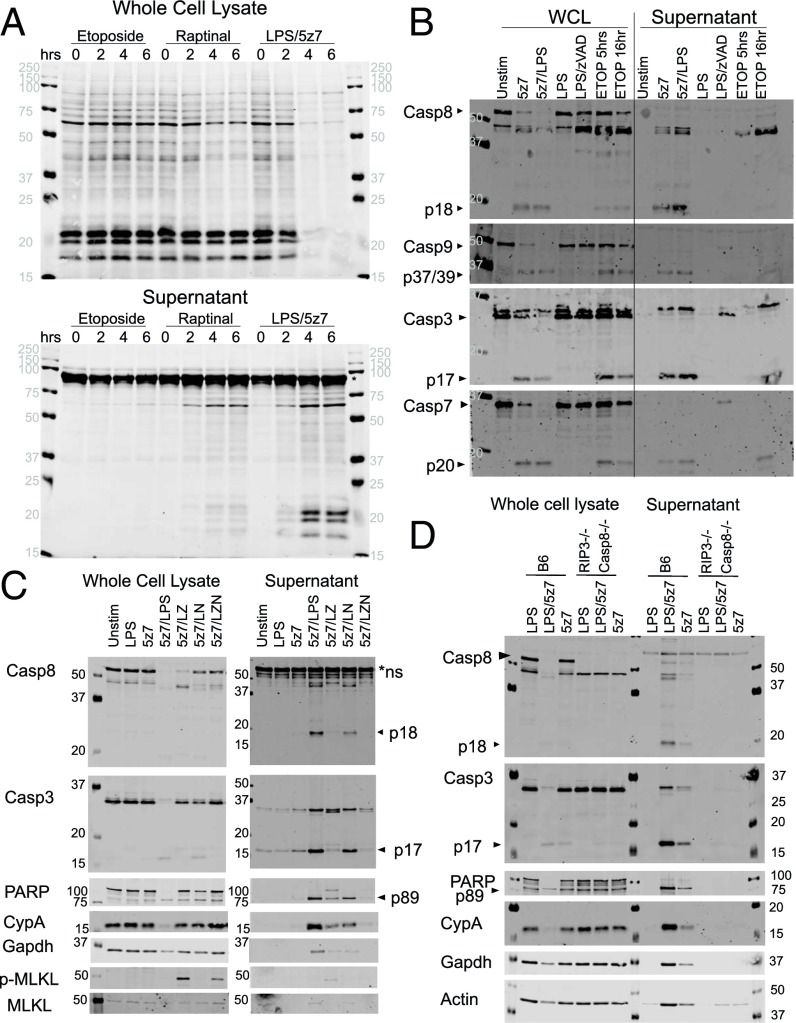
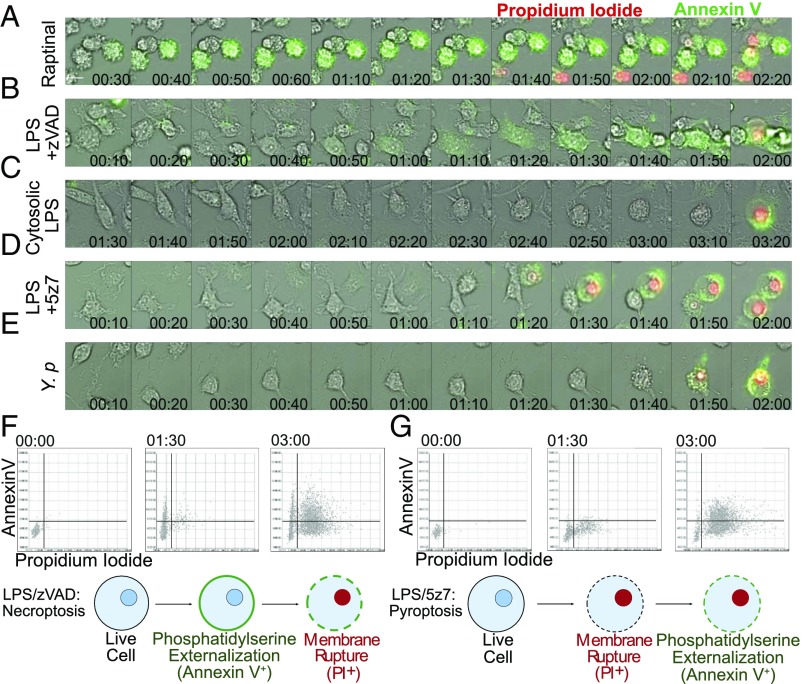
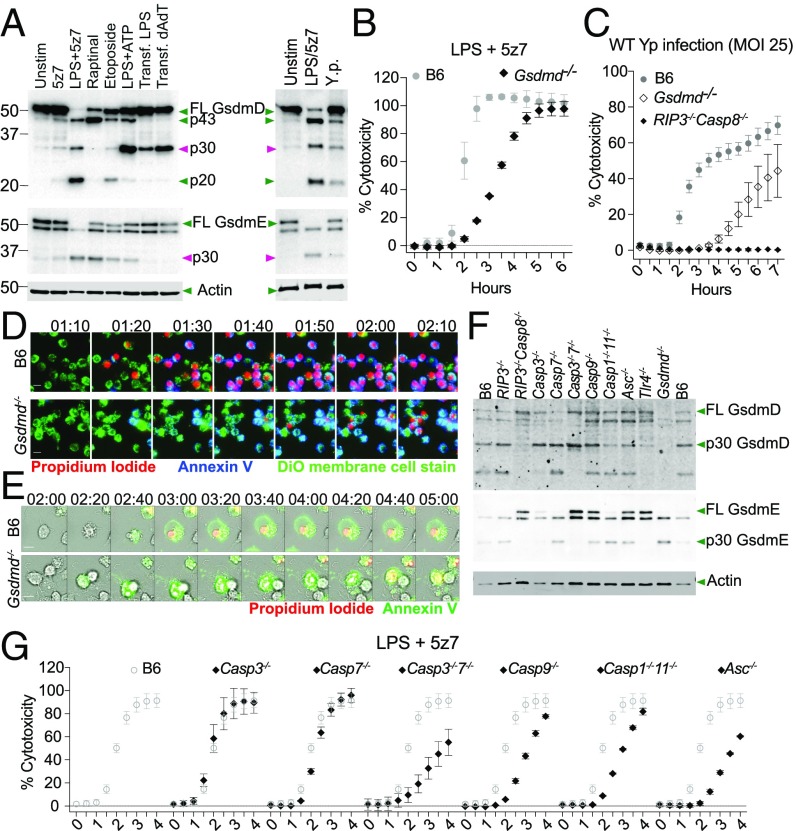
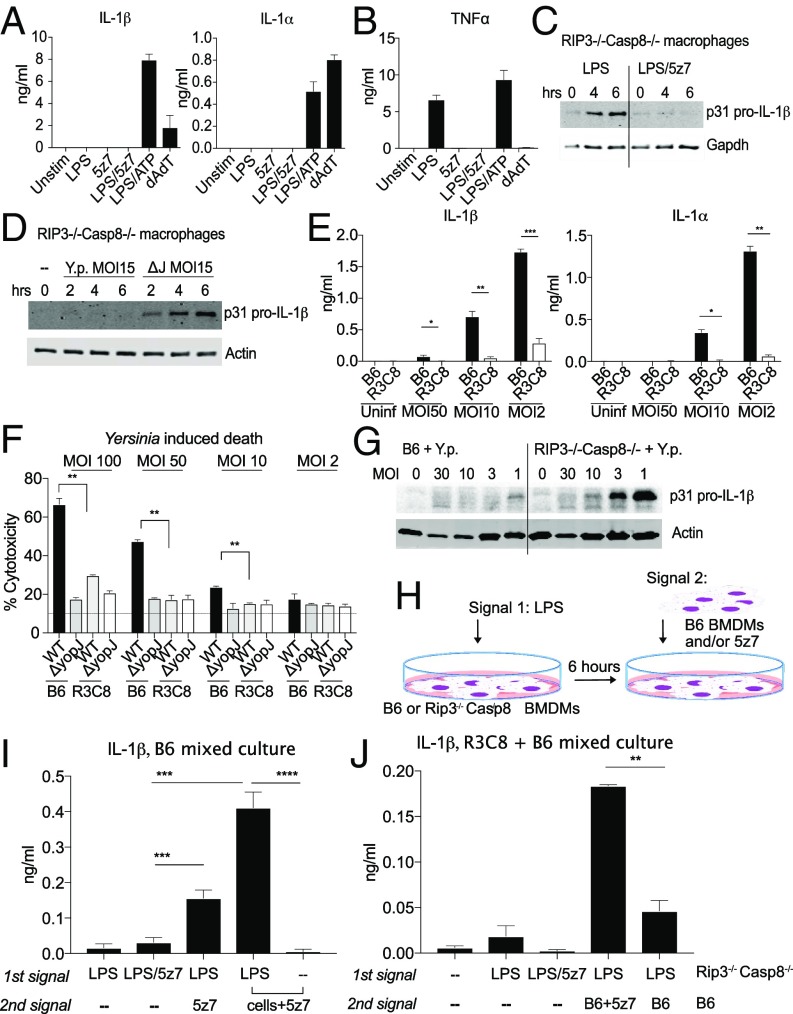
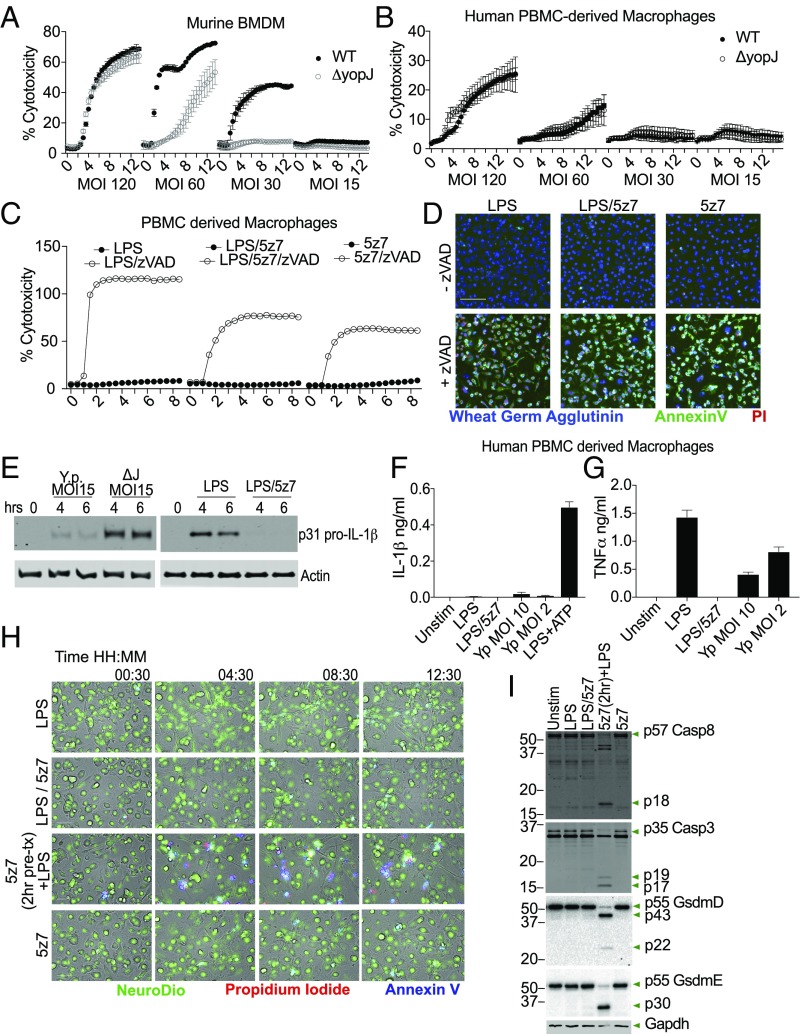
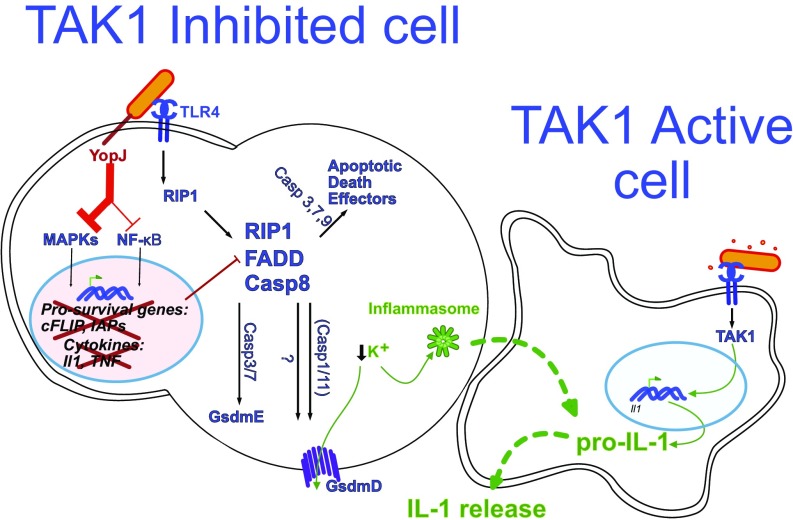
Cell death and inflammation are intimately linked during Yersinia infection. Pathogenic Yersinia inhibits the MAP kinase TGFβ-activated kinase 1 (TAK1) via the effector YopJ, thereby silencing cytokine expression while activating caspase-8-mediated cell death. Here, using Yersinia pseudotuberculosis in corroboration with costimulation of lipopolysaccharide and (5Z)-7-Oxozeaenol, a small-molecule inhibitor of TAK1, we show that caspase-8 activation during TAK1 inhibition results in cleavage of both gasdermin D (GSDMD) and gasdermin E (GSDME) in murine macrophages, resulting in pyroptosis. Loss of GsdmD delays membrane rupture, reverting the cell-death morphology to apoptosis. We found that the Yersinia-driven IL-1 response arises from asynchrony of macrophage death during bulk infections in which two cellular populations are required to provide signal 1 and signal 2 for IL-1α/β release. Furthermore, we found that human macrophages are resistant to YopJ-mediated pyroptosis, with dampened IL-1β production. Our results uncover a form of caspase-8-mediated pyroptosis and suggest a hypothesis for the increased sensitivity of humans to Yersinia infection compared with the rodent reservoir.
Keywords: TAK1; Yersinia; caspase-8; gasdermin; pyroptosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- O’Neill LA, Golenbock D, Bowie AG. The history of toll-like receptors–Redefining innate immunity. Nat Rev Immunol. 2013;13:453–460. - PubMed
-
- Park JM, et al. Signaling pathways and genes that inhibit pathogen-induced macrophage apoptosis–CREB and NF-kappaB as key regulators. Immunity. 2005;23:319–329. - PubMed
-
- Chen G, Goeddel DV. TNF-R1 signaling: A beautiful pathway. Science. 2002;296:1634–1635. - PubMed
-
- Kelliher MA, et al. The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity. 1998;8:297–303. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
