

The PDL1-inducible GTPase Arl4d controls T effector function by limiting IL-2 production
- PMID: 30382149
- PMCID: PMC6208435
- DOI: 10.1038/s41598-018-34522-4
The PDL1-inducible GTPase Arl4d controls T effector function by limiting IL-2 production
Abstract
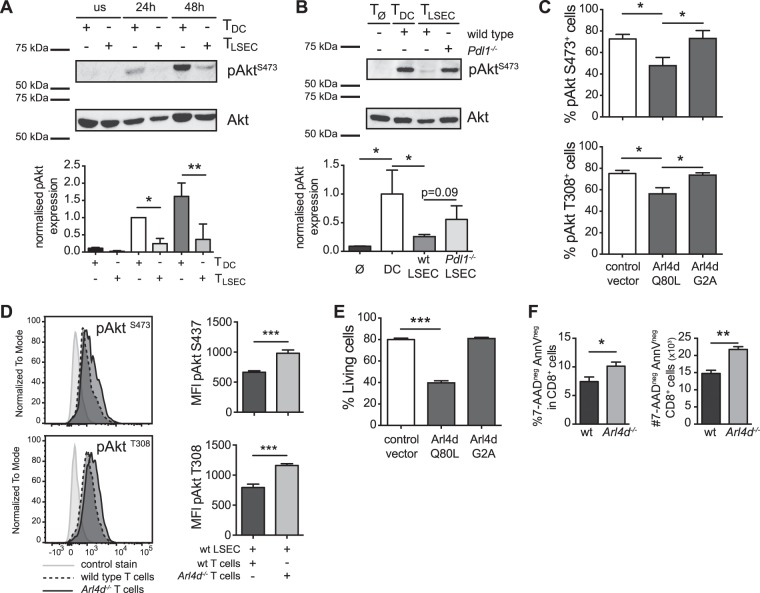
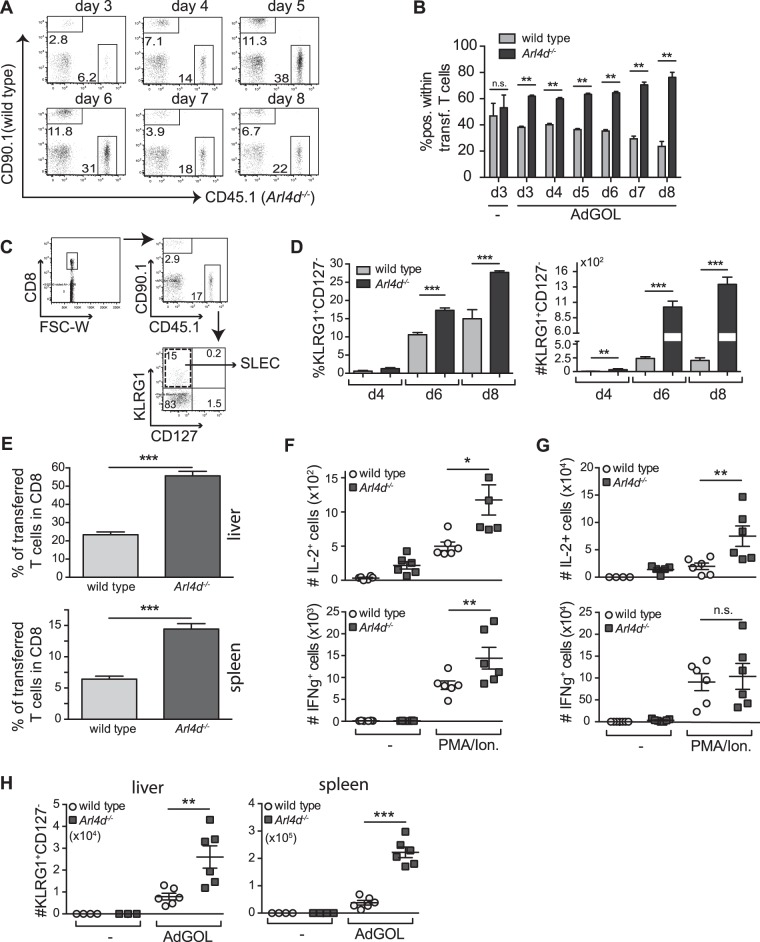
Interleukin-2 (IL-2) is a key regulator of adaptive immune responses but its regulation is incompletely understood. We previously found that PDL1-dependent signals were pivotal for liver sinusoidal endothelial cell-mediated priming of CD8 T cells, which have a strongly reduced capacity to produce IL-2. Here, we show that the expression of the ARF-like GTPase Arl4d is PD-L1-dependently induced in such LSEC-primed T cells, and is associated with reduced IL-2 secretion and Akt phosphorylation. Conversely, Arl4d-deficient T cells overproduced IL-2 upon stimulation. Arl4d-deficiency in CD8 T cells also enhanced their expansion and effector function during viral infection in vivo. Consistent with their increased IL-2 production, Arl4d-deficient T cells showed enhanced development into KLRG1+CD127- short-lived effector cells (SLEC), which is dependent on IL-2 availability. Thus, our data reveal a PD-L1-dependent regulatory circuitry that involves the induction of Arl4d for limiting IL-2 production in T cells.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
