

Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury
- PMID: 30382196
- PMCID: PMC6268129
- DOI: 10.1038/s41593-018-0262-6
Electrical spinal cord stimulation must preserve proprioception to enable locomotion in humans with spinal cord injury
Abstract
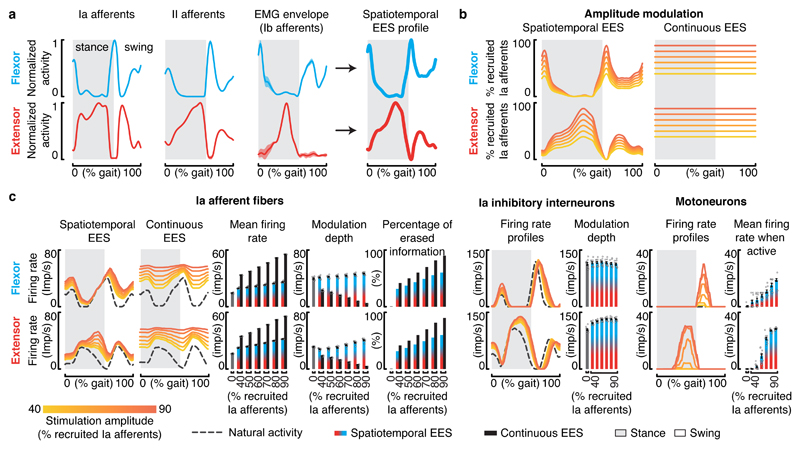
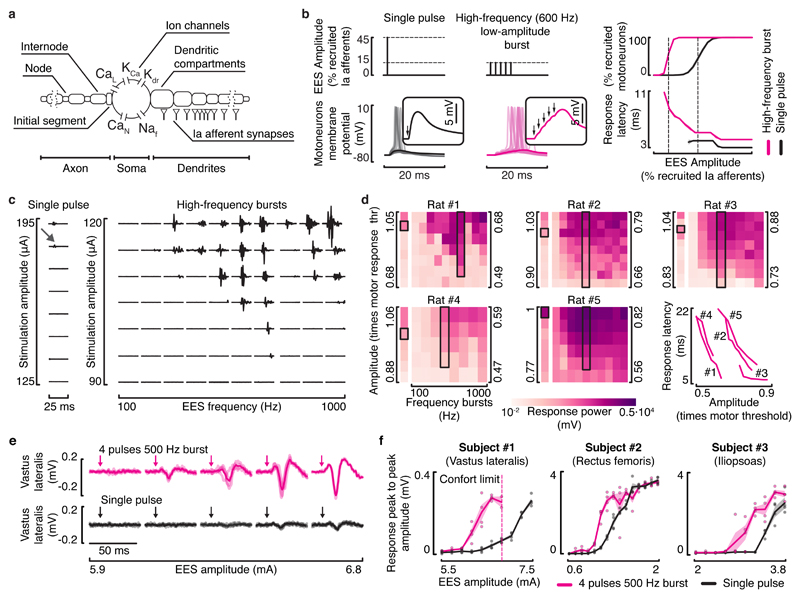
Epidural electrical stimulation (EES) of the spinal cord restores locomotion in animal models of spinal cord injury but is less effective in humans. Here we hypothesized that this interspecies discrepancy is due to interference between EES and proprioceptive information in humans. Computational simulations and preclinical and clinical experiments reveal that EES blocks a significant amount of proprioceptive input in humans, but not in rats. This transient deafferentation prevents modulation of reciprocal inhibitory networks involved in locomotion and reduces or abolishes the conscious perception of leg position. Consequently, continuous EES can only facilitate locomotion within a narrow range of stimulation parameters and is unable to provide meaningful locomotor improvements in humans without rehabilitation. Simulations showed that burst stimulation and spatiotemporal stimulation profiles mitigate the cancellation of proprioceptive information, enabling robust control over motor neuron activity. This demonstrates the importance of stimulation protocols that preserve proprioceptive information to facilitate walking with EES.
Conflict of interest statement
G.C. and S.M. are founders and shareholders of GTXmedical SA, a company developing neuroprosthetic systems in direct relationship with the present work. E.F., M.C., G.C. and S.M. hold several patents related to electrical spinal cord stimulation.
Figures

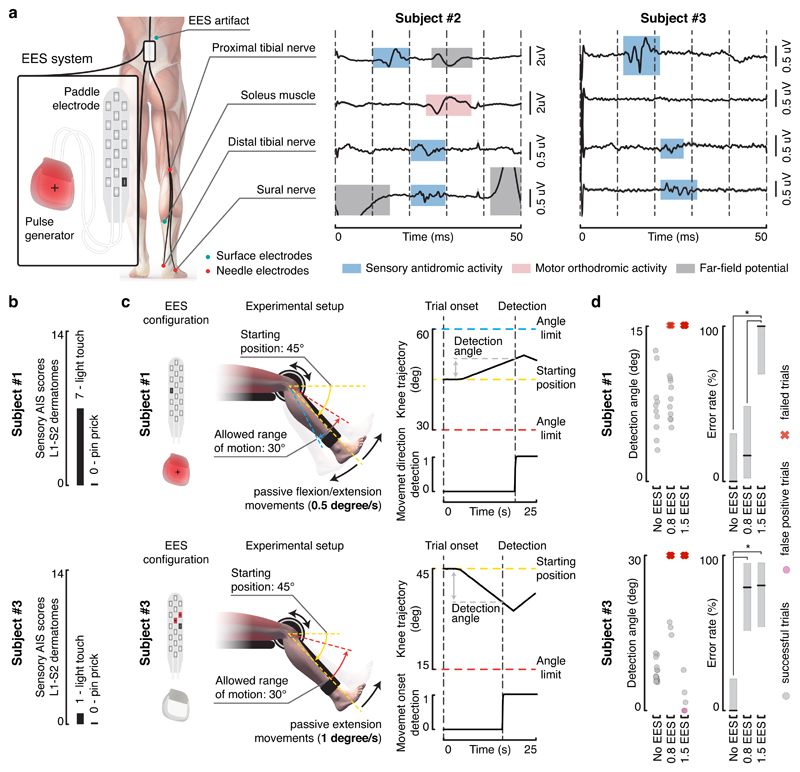

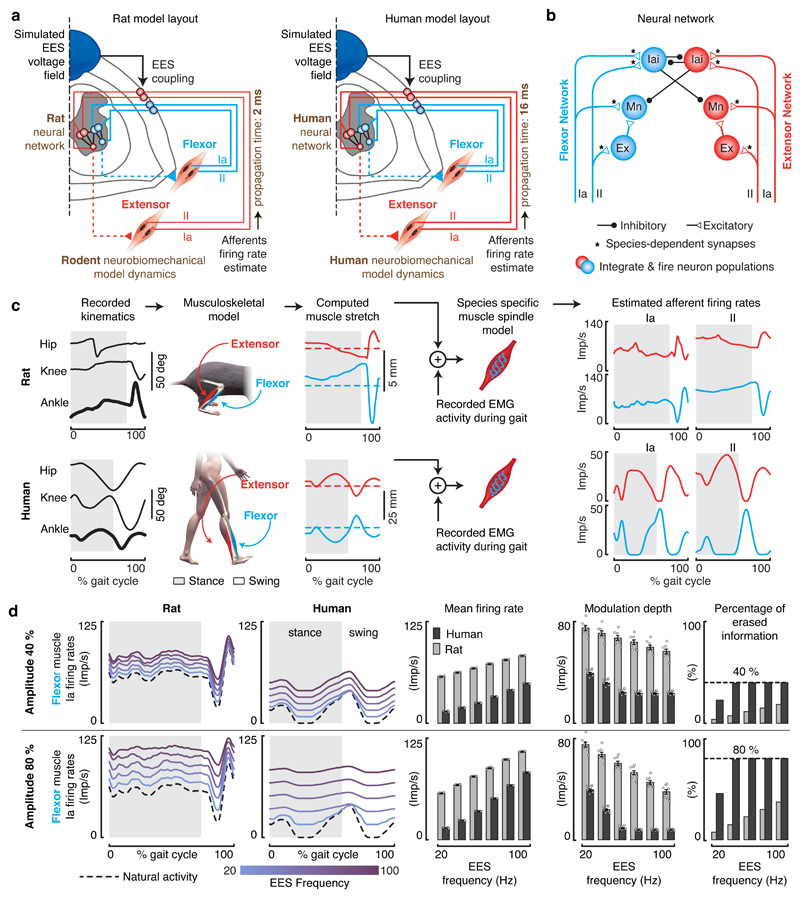
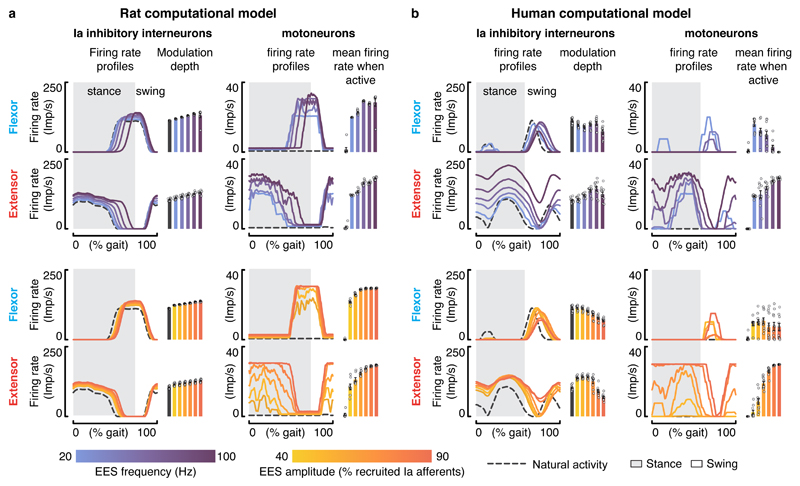
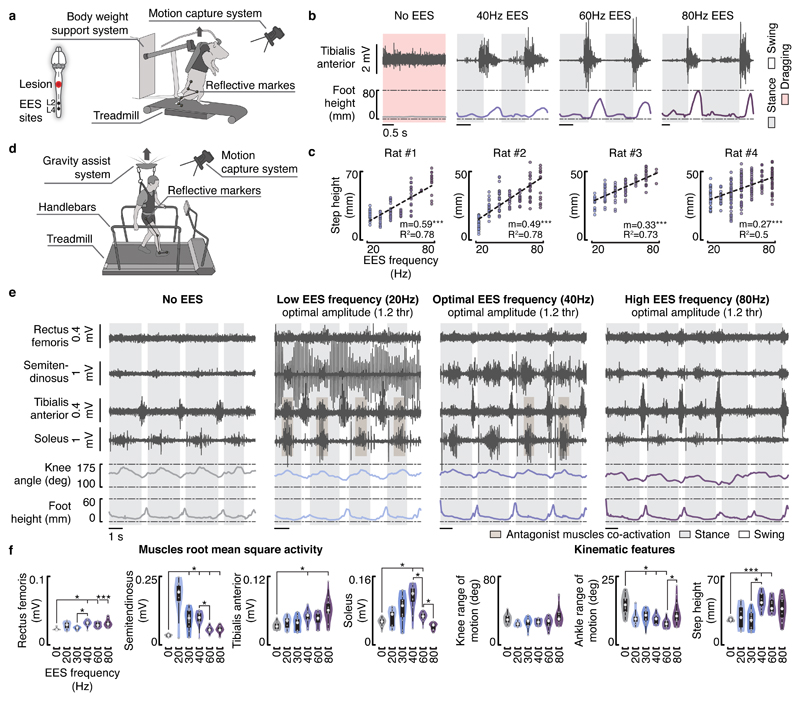
Comment in
-
A giant step for spinal cord injury research.Nat Neurosci. 2018 Dec;21(12):1647-1648. doi: 10.1038/s41593-018-0264-4. Nat Neurosci. 2018. PMID: 30382195 No abstract available.
-
Restoring walking.Nat Rev Neurosci. 2019 Jan;20(1):1. doi: 10.1038/s41583-018-0109-x. Nat Rev Neurosci. 2019. PMID: 30518965 No abstract available.
References
-
- Van den Brand R, et al. Restoring Voluntary Control of Locomotion after Paralyzing Spinal Cord Injury. Science. 2012;336:1182–1185. - PubMed
-
- Wenger N, et al. Closed-loop neuromodulation of spinal sensorimotor circuits controls refined locomotion after complete spinal cord injury. Science Translational Medicine. 2014;6:255ra133–255ra133. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
