

Hemoglobin oxidation-dependent reactions promote interactions with band 3 and oxidative changes in sickle cell-derived microparticles
- PMID: 30385713
- PMCID: PMC6238743
- DOI: 10.1172/jci.insight.120451
Hemoglobin oxidation-dependent reactions promote interactions with band 3 and oxidative changes in sickle cell-derived microparticles
Abstract
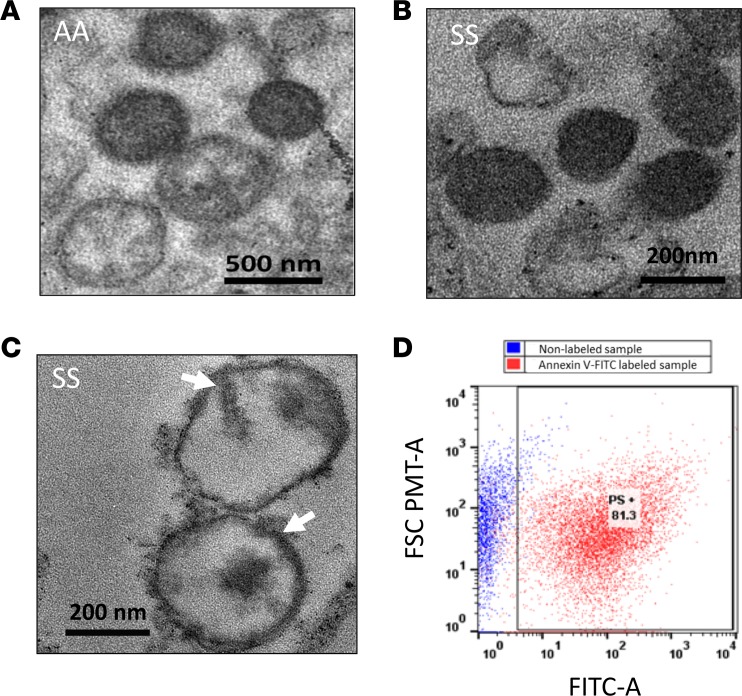
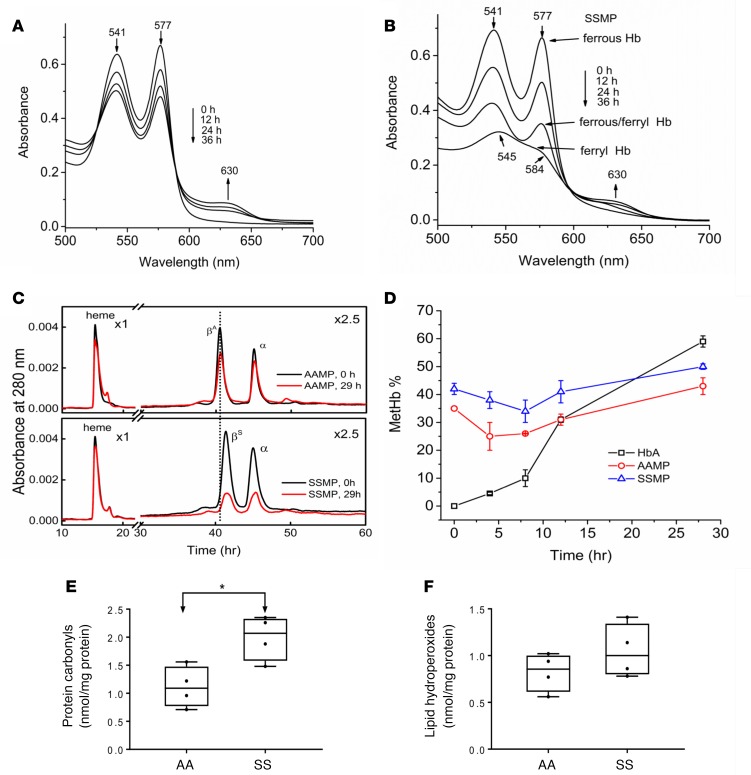
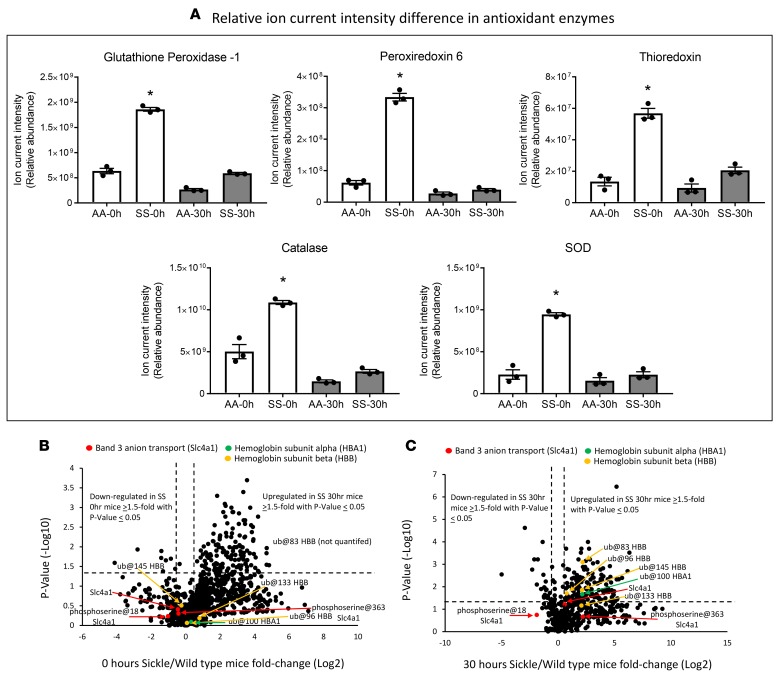
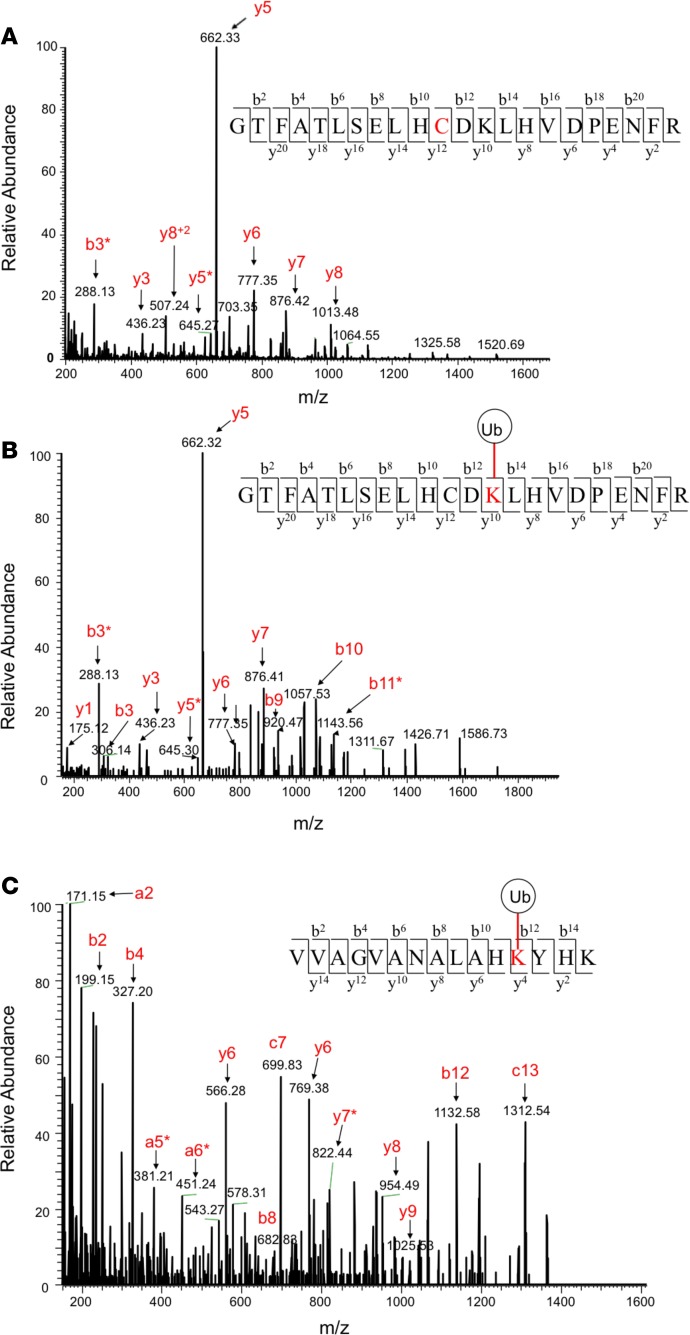
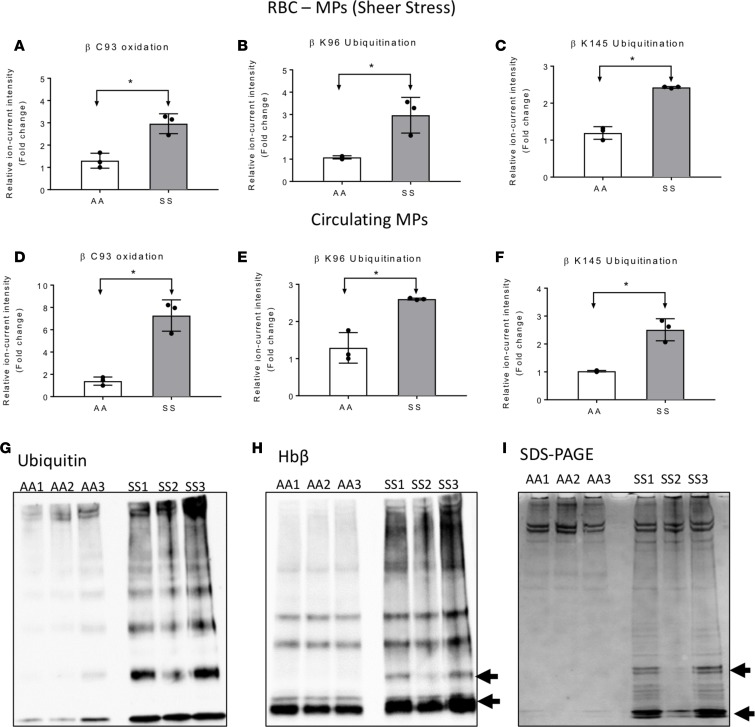
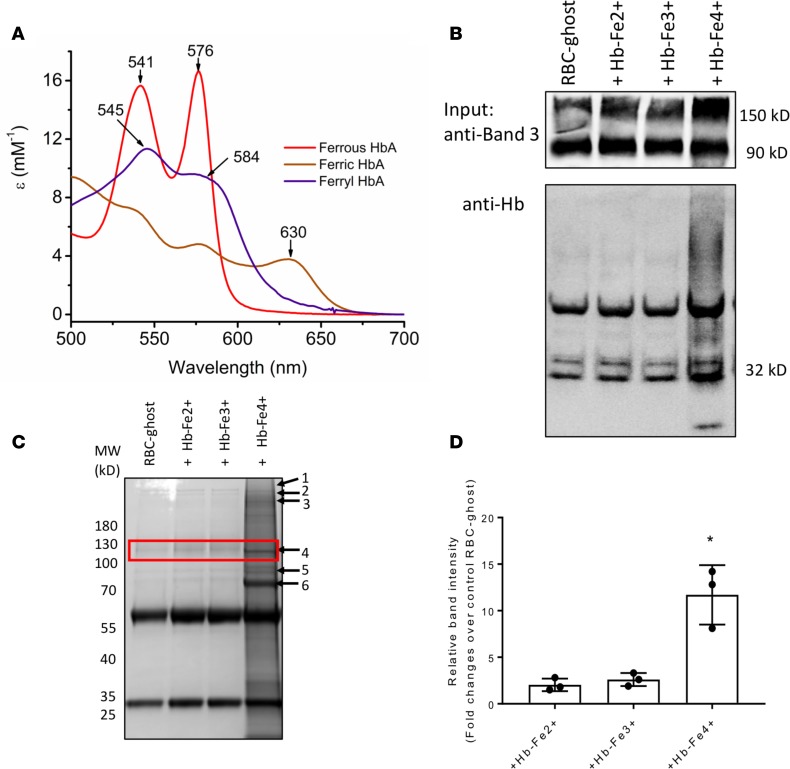
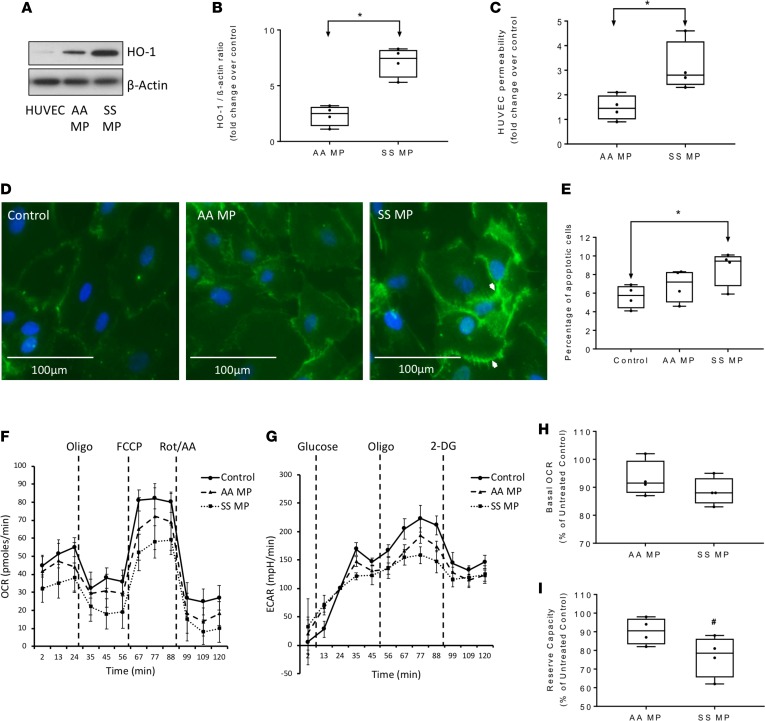
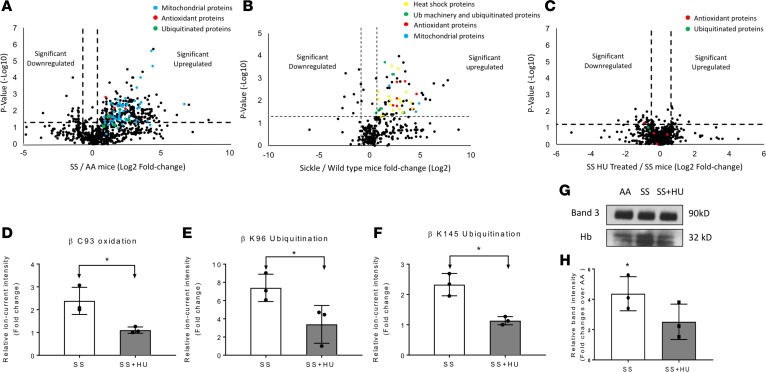
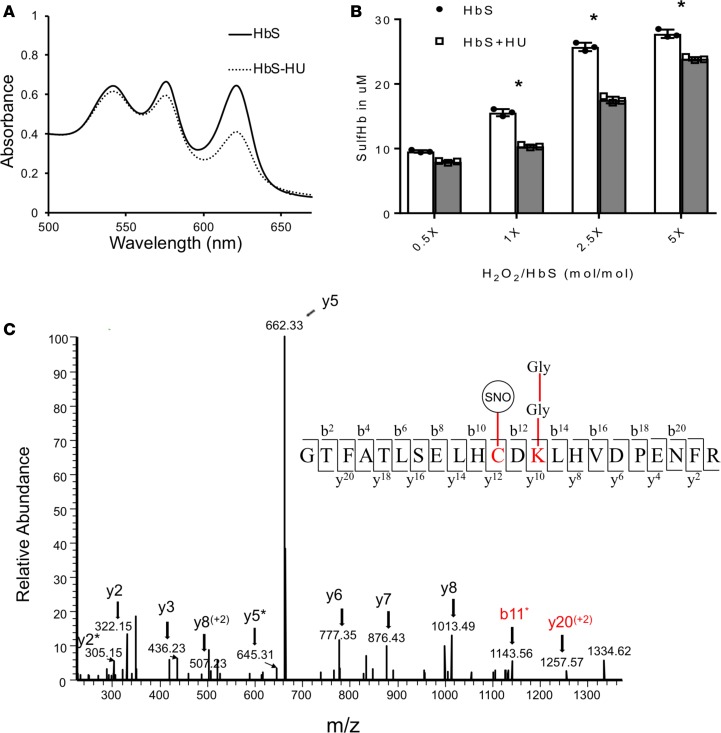
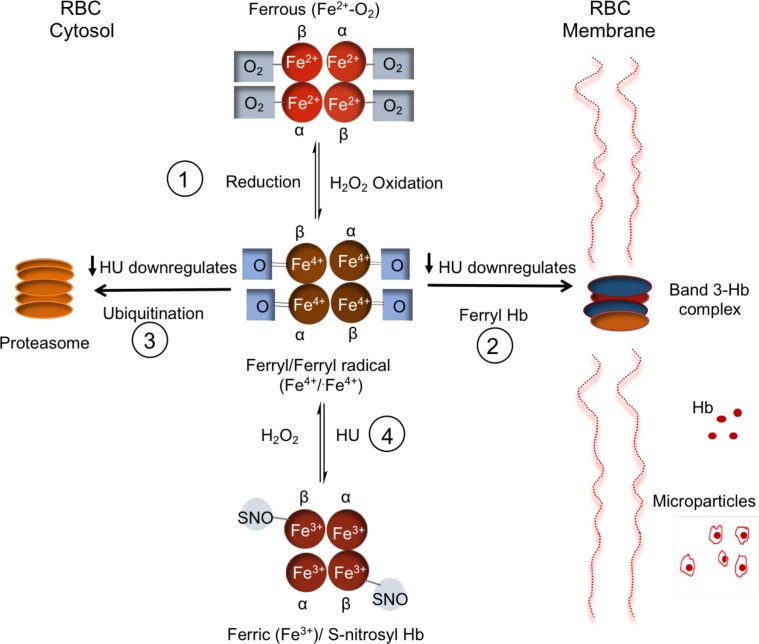
The contribution of intracellular hemoglobin (Hb) oxidation to RBC-derived microparticle (MP) formation is poorly defined in sickle cell disease (SCD). Here we report that sickle Hb (HbS) oxidation, coupled with changes in cytosolic antioxidative proteins, is associated with membrane alterations and MP formation in homozygous Townes-sickle cell (Townes-SS) mice. Photometric and proteomic analyses confirmed the presence of high levels of Hb oxidation intermediates (ferric/ferryl) and consequent β-globin posttranslational modifications, including the irreversible oxidation of βCys93 and the ubiquitination of βLys96 and βLys145. This is the first report to our knowledge to link the UPS (via ubiquitinated Hb and other proteins) to oxidative stress. Ferryl Hb also induced complex formation with band 3 and RBC membrane proteins. Incubation of Townes-SS MPs with human endothelial cells caused greater loss of monolayer integrity, apoptotic activation, heme oxygenase-1 induction, and concomitant bioenergetic imbalance compared with control Townes-AA MPs. MPs obtained from Townes-SS mice treated with hydroxyurea produced fewer posttranslational Hb modifications. In vitro, hydroxyurea reduced the levels of ferryl Hb and shielded its target residue, βCys93, by a process of S-nitrosylation. These mechanistic analyses suggest potential antioxidative therapeutic modalities that may interrupt MP heme-mediated pathophysiology in SCD patients.
Keywords: Cardiovascular disease; Hematology; Proteomics; Ubiquitin-proteosome system; Vascular Biology.
Conflict of interest statement
Figures
Comment in
-
Posttranslational modifications of sickle hemoglobin in microparticles may promote injury.Kidney Int. 2019 Jun;95(6):1289-1291. doi: 10.1016/j.kint.2019.02.023. Kidney Int. 2019. PMID: 31122703 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
