

Active Human Complement Reduces the Zika Virus Load via Formation of the Membrane-Attack Complex
- PMID: 30386325
- PMCID: PMC6199351
- DOI: 10.3389/fimmu.2018.02177
Active Human Complement Reduces the Zika Virus Load via Formation of the Membrane-Attack Complex
Abstract
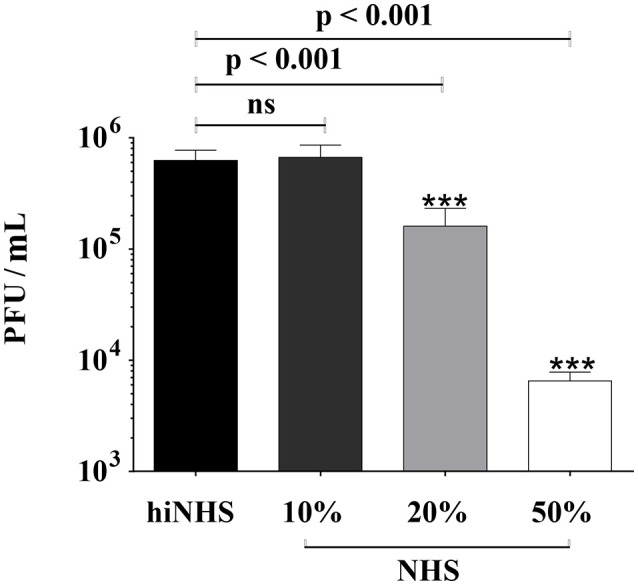
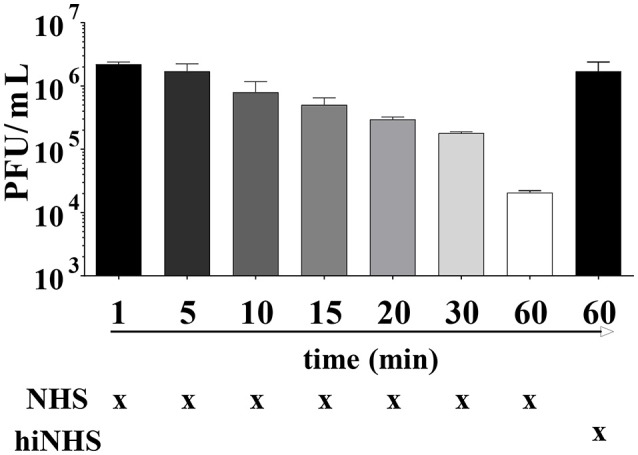
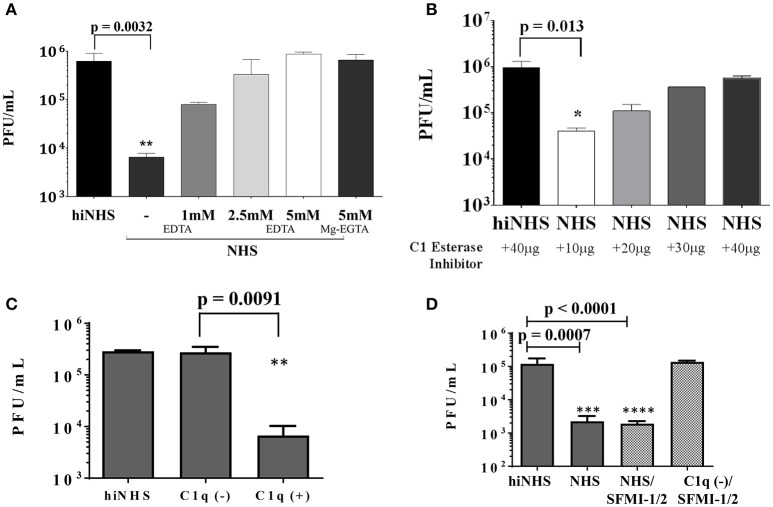
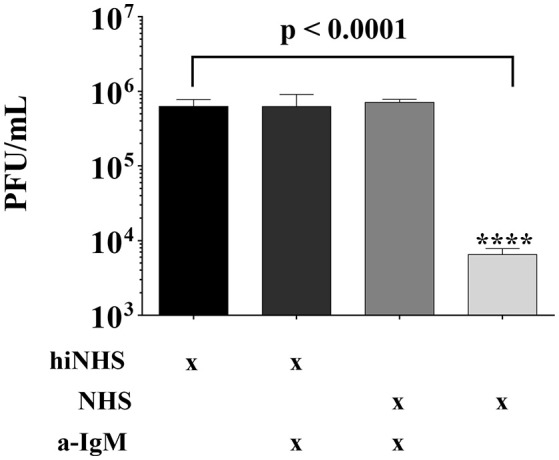
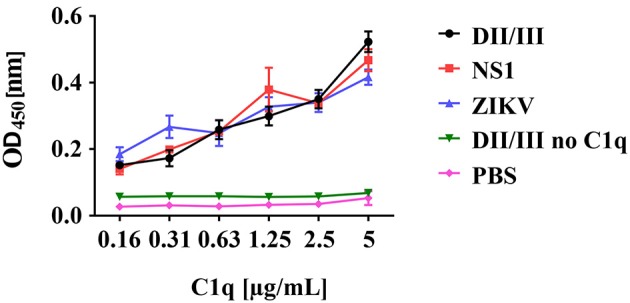
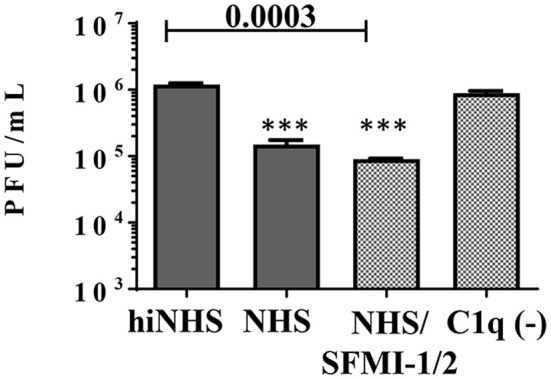
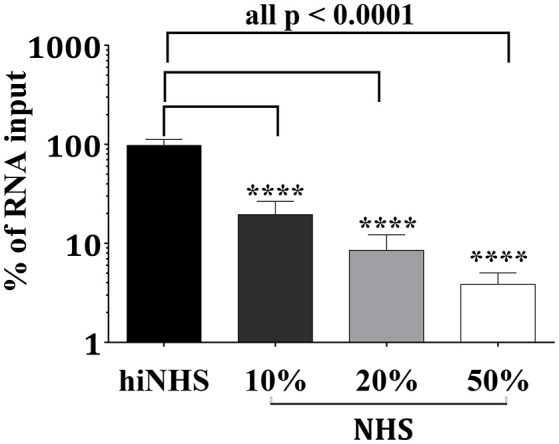
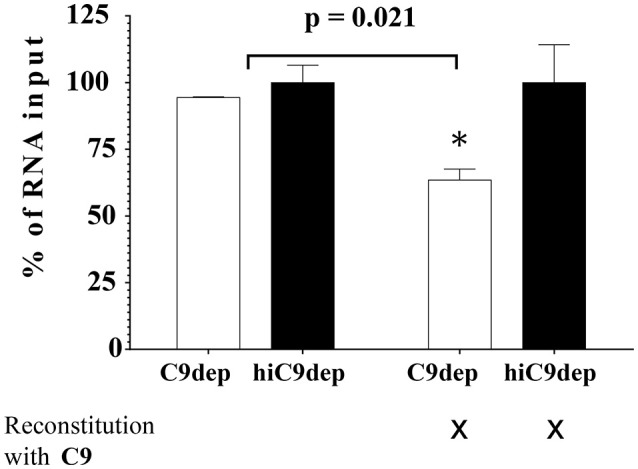
Although neglected in the past, the interest on Zika virus (ZIKV) raised dramatically in the last several years. The rapid spread of the virus in Latin America and the association of the infection with microcephaly in newborns or Guillain-Barré Syndrome in adults prompted the WHO to declare the ZIKV epidemic to be an international public health emergency in 2016. As the virus gained only limited attention in the past, investigations on interactions of ZIKV with human complement are limited. This prompted us to investigate the stability of the virus to human complement. At low serum concentrations (10%) which refers to complement concentrations found on mucosal surfaces, the virus was relatively stable at 37°C, while at high complement levels (50% serum concentration) ZIKV titers were dramatically reduced, although the virus remained infectious for about 4-5 min under these conditions. The classical pathway was identified as the main actor of complement activation driven by IgM antibodies. In addition, direct binding of C1q to both envelope and NS1 proteins was observed. Formation of the MAC on the viral surface and thus complement-mediated lysis and not opsonization seems to be essential for the reduction of viral titers.
Keywords: C1q; IgM; MAC; Zika virus; complement; lysis.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous