

Exon Junction Complex Shapes the Transcriptome by Repressing Recursive Splicing
- PMID: 30388411
- PMCID: PMC6224609
- DOI: 10.1016/j.molcel.2018.09.033
Exon Junction Complex Shapes the Transcriptome by Repressing Recursive Splicing
Abstract
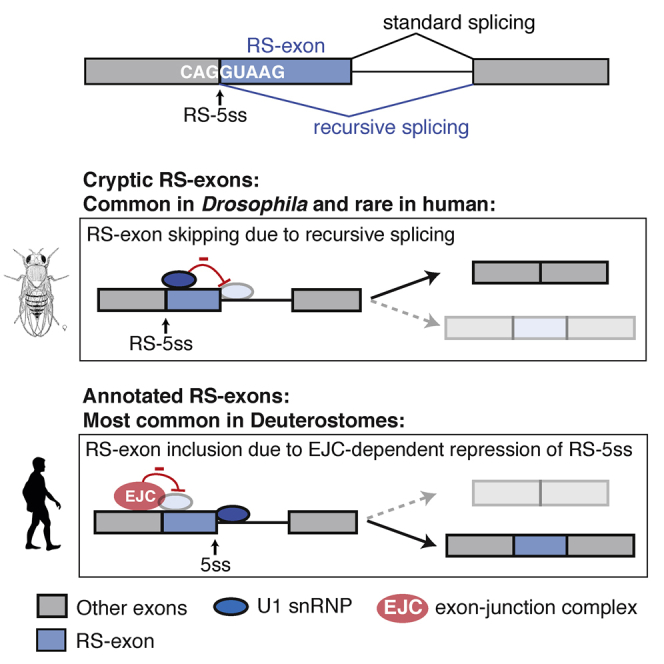
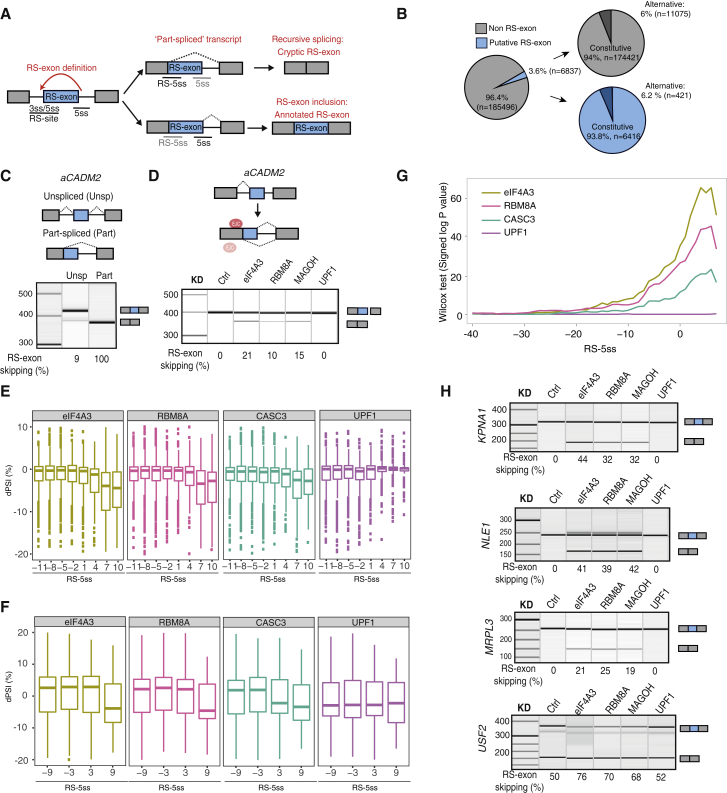
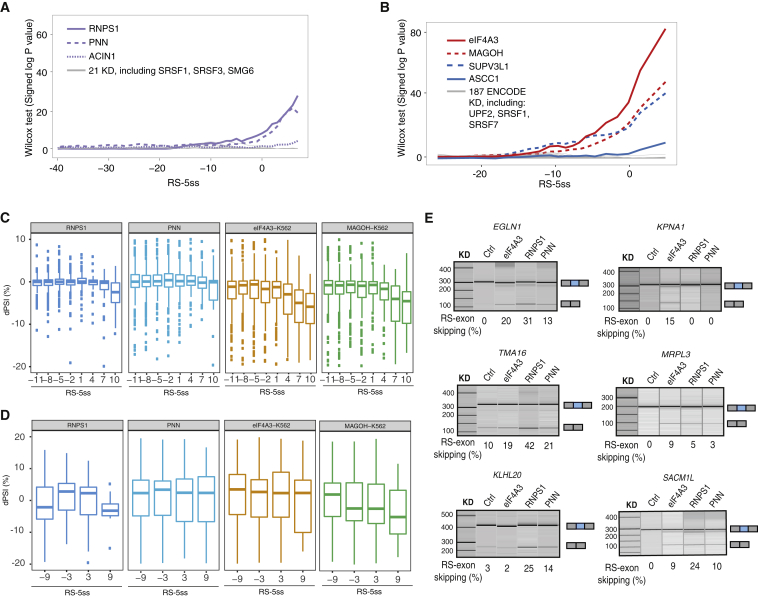
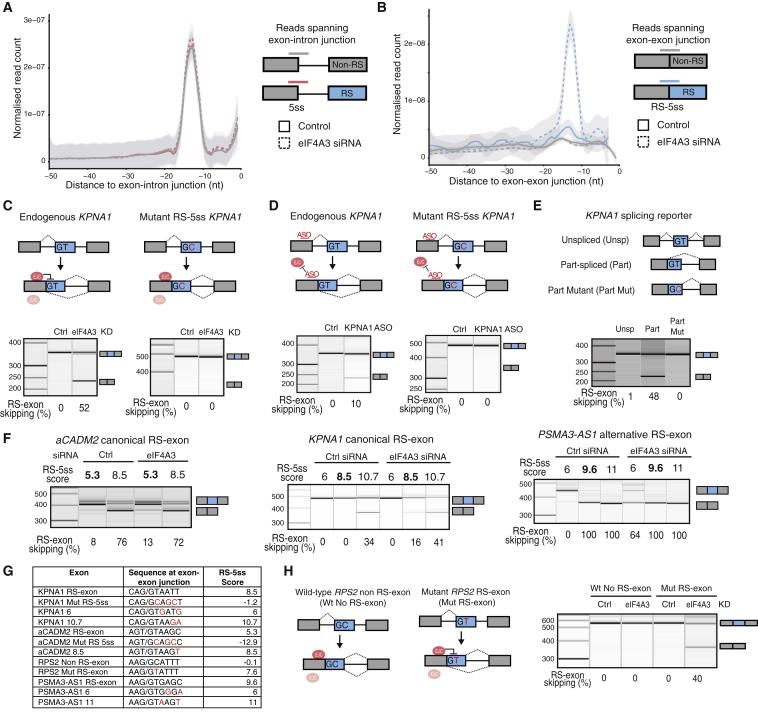
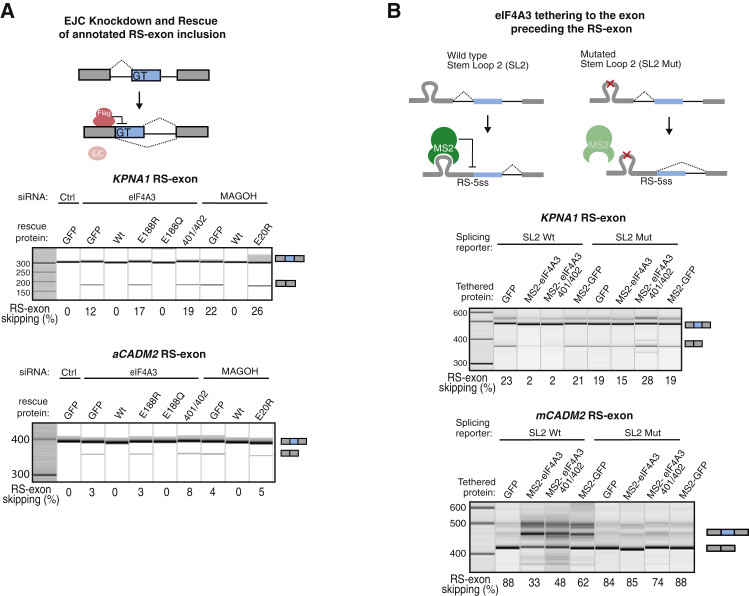
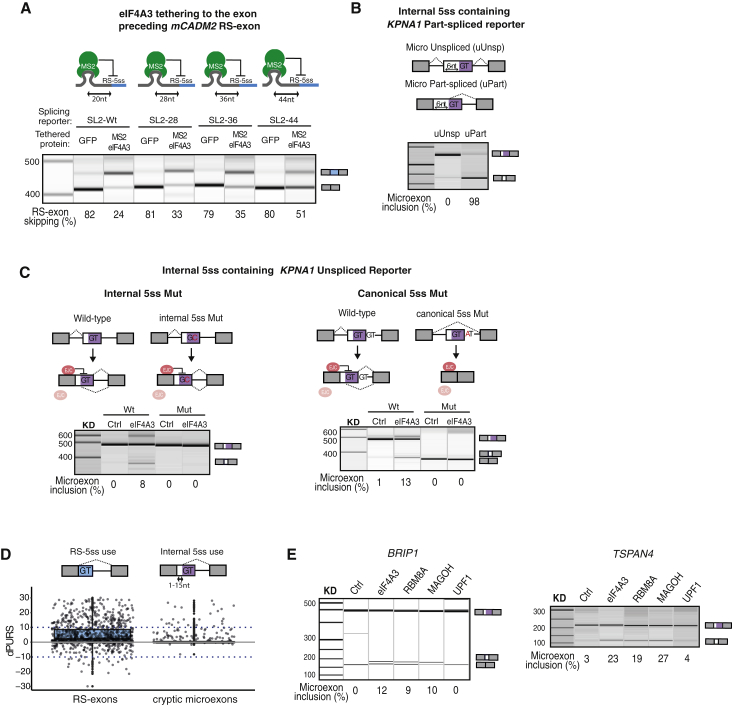
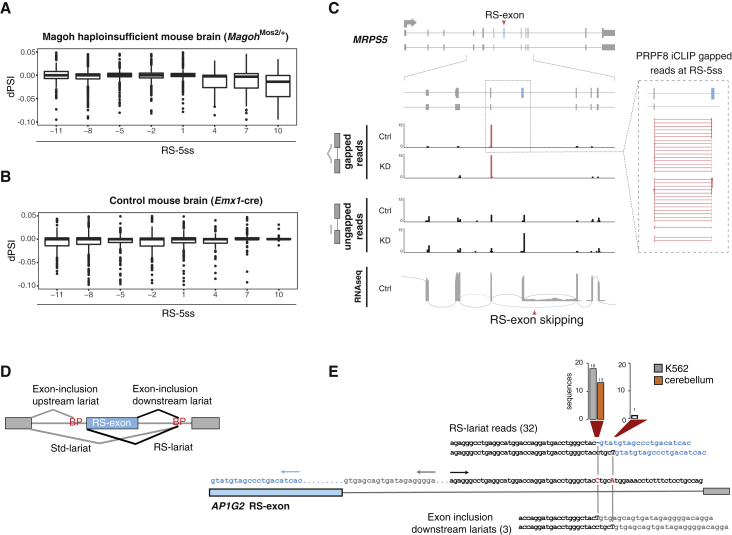
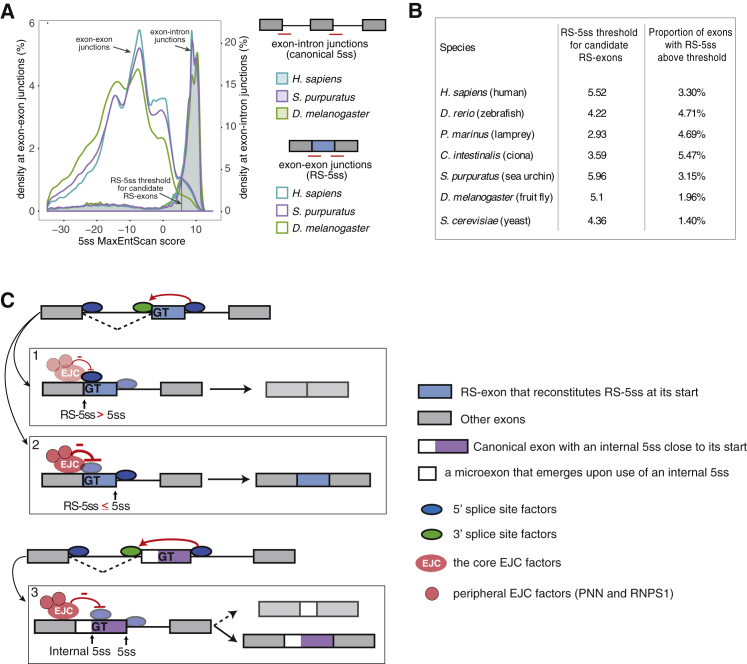
Recursive splicing (RS) starts by defining an "RS-exon," which is then spliced to the preceding exon, thus creating a recursive 5' splice site (RS-5ss). Previous studies focused on cryptic RS-exons, and now we find that the exon junction complex (EJC) represses RS of hundreds of annotated, mainly constitutive RS-exons. The core EJC factors, and the peripheral factors PNN and RNPS1, maintain RS-exon inclusion by repressing spliceosomal assembly on RS-5ss. The EJC also blocks 5ss located near exon-exon junctions, thus repressing inclusion of cryptic microexons. The prevalence of annotated RS-exons is high in deuterostomes, while the cryptic RS-exons are more prevalent in Drosophila, where EJC appears less capable of repressing RS. Notably, incomplete repression of RS also contributes to physiological alternative splicing of several human RS-exons. Finally, haploinsufficiency of the EJC factor Magoh in mice is associated with skipping of RS-exons in the brain, with relevance to the microcephaly phenotype and human diseases.
Keywords: RS exon; alternative splicing mechanisms; evolution; exon junction complex; gene expression; microcephaly; microexon; neurodevelopmental disorders; recursive splicing.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
The Exon Junction Complex: A Multitasking Guardian of the Transcriptome.Mol Cell. 2018 Dec 6;72(5):799-801. doi: 10.1016/j.molcel.2018.11.030. Mol Cell. 2018. PMID: 30526869 Free PMC article.
References
-
- Ashton-Beaucage D., Udell C.M., Lavoie H., Baril C., Lefrançois M., Chagnon P., Gendron P., Caron-Lizotte O., Bonneil E., Thibault P., Therrien M. The exon junction complex controls the splicing of MAPK and other long intron-containing transcripts in Drosophila. Cell. 2010;143:251–262. - PubMed
-
- Ballut L., Marchadier B., Baguet A., Tomasetto C., Séraphin B., Le Hir H. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat. Struct. Mol. Biol. 2005;12:861–869. - PubMed
-
- Bartkowska K., Tepper B., Turlejski K., Djavadian R.L. Roles of the exon junction complex components in the central nervous system: a mini review. Rev. Neurosci. 2018 Published online May 23, 2018. - PubMed
-
- Boehm V., Britto-Borges T., Steckelberg A.-L., Singh K.K., Gerbracht J.V., Gueney E., Blazquez L., Altmueller J., Dieterich C., Gehring N.H. Exon junction complexes suppress spurious splice sites to safeguard transcriptome integrity. Mol. Cell. 2018;72:482–495. this issue. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
