

Two to Tango: Dialog between Immunity and Stem Cells in Health and Disease
- PMID: 30388451
- PMCID: PMC6294328
- DOI: 10.1016/j.cell.2018.08.071
Two to Tango: Dialog between Immunity and Stem Cells in Health and Disease
Abstract
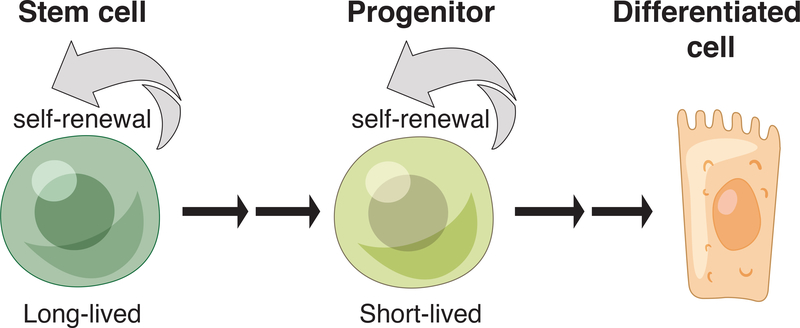
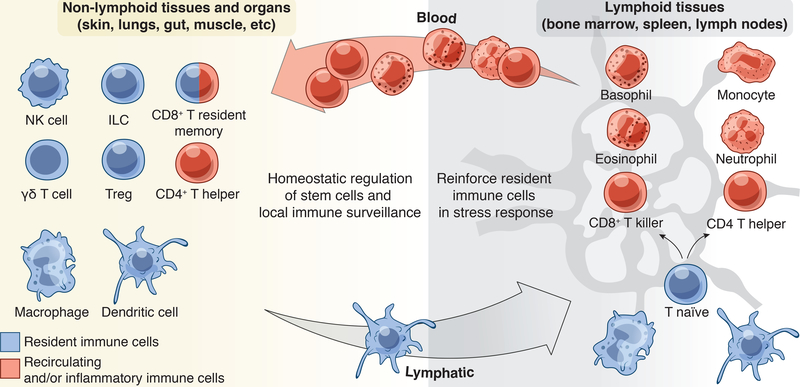
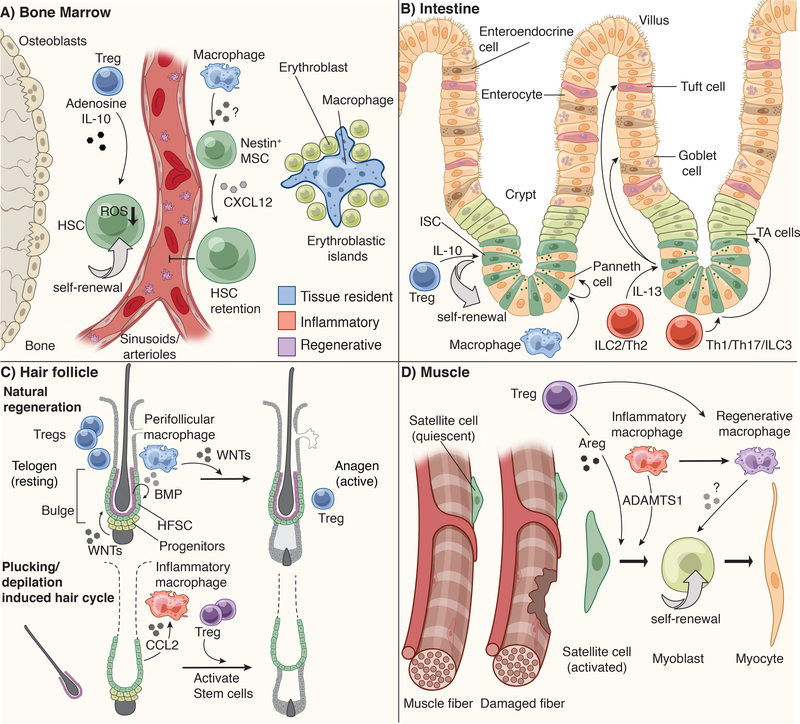
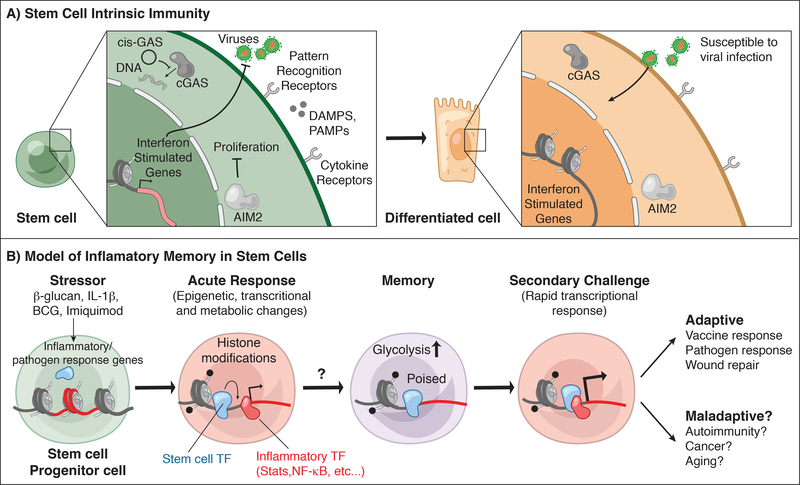
Stem cells regenerate tissues in homeostasis and under stress. By taking cues from their microenvironment or "niche," they smoothly transition between these states. Immune cells have surfaced as prominent members of stem cell niches across the body. Here, we draw parallels between different stem cell niches to explore the context-specific interactions that stem cells have with tissue-resident and recruited immune cells. We also highlight stem cells' innate ability to sense and respond to stress and the enduring memory that forms from such encounters. This fascinating crosstalk holds great promise for novel therapies in inflammatory diseases and regenerative medicine.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
References
-
- Alvarado AG, and Lathia JD (2016). Taking a Toll on Self-Renewal: TLR-Mediated Innate Immune Signaling in Stem Cells. Trends in Neurosciences 39, 463–471. - PubMed
-
- Aparicio-Domingo P, Romera-Hernandez M, Karrich JJ, Cornelissen F, Papazian N, Lindenbergh-Kortleve DJ, Butler JA, Boon L, Coles MC, Samsom JN, et al. (2015). Type 3 innate lymphoid cells maintain intestinal epithelial stem cells after tissue damage. The Journal of Experimental Medicine 212, 1783–1791. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
