

The kinetochore module Okp1CENP-Q/Ame1CENP-U is a reader for N-terminal modifications on the centromeric histone Cse4CENP-A
- PMID: 30389668
- PMCID: PMC6315295
- DOI: 10.15252/embj.201898991
The kinetochore module Okp1CENP-Q/Ame1CENP-U is a reader for N-terminal modifications on the centromeric histone Cse4CENP-A
Abstract
Kinetochores are supramolecular assemblies that link centromeres to microtubules for sister chromatid segregation in mitosis. For this, the inner kinetochore CCAN/Ctf19 complex binds to centromeric chromatin containing the histone variant CENP-A, but whether the interaction of kinetochore components to centromeric nucleosomes is regulated by posttranslational modifications is unknown. Here, we investigated how methylation of arginine 37 (R37Me) and acetylation of lysine 49 (K49Ac) on the CENP-A homolog Cse4 from Saccharomyces cerevisiae regulate molecular interactions at the inner kinetochore. Importantly, we found that the Cse4 N-terminus binds with high affinity to the Ctf19 complex subassembly Okp1/Ame1 (CENP-Q/CENP-U in higher eukaryotes), and that this interaction is inhibited by R37Me and K49Ac modification on Cse4. In vivo defects in cse4-R37A were suppressed by mutations in OKP1 and AME1, and biochemical analysis of a mutant version of Okp1 showed increased affinity for Cse4. Altogether, our results demonstrate that the Okp1/Ame1 heterodimer is a reader module for posttranslational modifications on Cse4, thereby targeting the yeast CCAN complex to centromeric chromatin.
Keywords: Ame1; Gcn5; Okp1; centromere; posttranslational modification.
© 2018 The Authors.
Figures
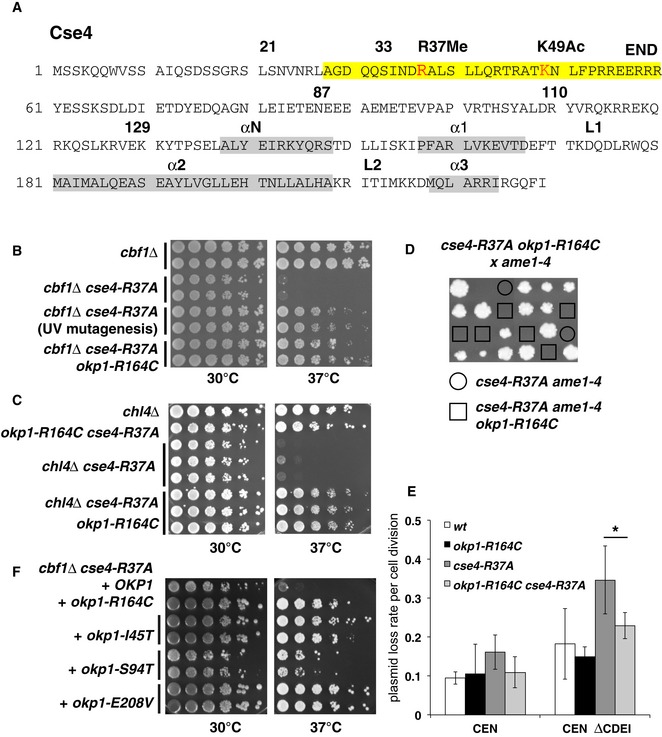
Overview of the amino acid sequence of Cse4. R37Me and K49Ac sites are part of the essential N‐terminal domain (END, aa 28–60, yellow) and are indicated in red. The localization of α‐helices in the histone fold domain is shown in grey. Amino acid residues that are relevant for this study are indicated with numbers.
okp1‐R164C suppressed the temperature‐sensitive growth defect of cbf1∆ cse4‐R37A. Serial dilutions of strains with the indicated genotypes were spotted on full medium and grown for 3 days at 30 or 37°C. The original okp1‐R164C isolate from the suppressor screen is indicated as “UV mutagenesis”.
okp1‐R164C suppressed the growth defect of cse4‐R37A with chl4∆. Representation as in (B).
okp1‐R164C was unable to suppress the lethality of cse4‐R37A with ame1‐4. Tetrad dissection of a genetic cross between a cse4‐R37A okp1‐R164C and an ame1‐4 strain is shown. The four spores from individual asci are aligned in vertical rows.
okp1‐R164C suppressed the maintenance defect of cse4‐R37A for plasmids lacking the CDEI sequence of CEN6 (CEN ∆CDEI, at 37°C). Error bars give SD of at least three independent transformants. *P = 0.03.
The okp1 mutations I45T, S94T and E208V suppressed the temperature‐sensitive growth defect of cbf1∆ cse4‐R37A. Strains with the indicated okp1 alleles on a plasmid (derivatives of AEY5584) are shown as in (B).
- A
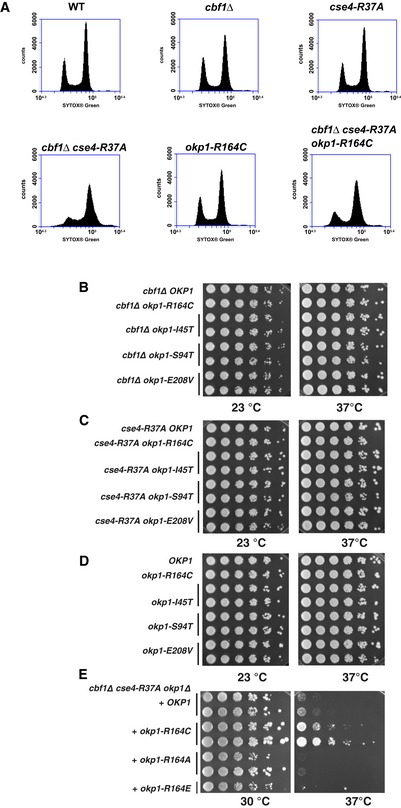
WT (AEY4), cbf1Δ (AEY4816), cse4‐R37A (AEY4965), cbf1Δ cse4‐R37A (AEY4985), okp1‐R164C (AEY5594) and cbf1Δ cse4‐R37A okp1‐R164C (AEY5974) cells were grown to early logarithmic phase at 23°C and shifted to 37°C for 5 h. DNA content of cells stained with SYTOX Green was measured by FACS analysis.
- B–D
Characterization of okp1 alleles. Alleles of okp1 that suppress cbf1∆ cse4‐R37A defects had no defect with cbf1∆ (B), cse4‐R37A (C) or in a wild‐type background (D). Strains carrying the indicated alleles were serially diluted and grown on full medium at the indicated temperatures for 3 days.
- E
Mutation Okp1‐R164 to alanine or glutamate did not suppress the temperature‐sensitive defect of cbf1∆ cse4‐R37A.
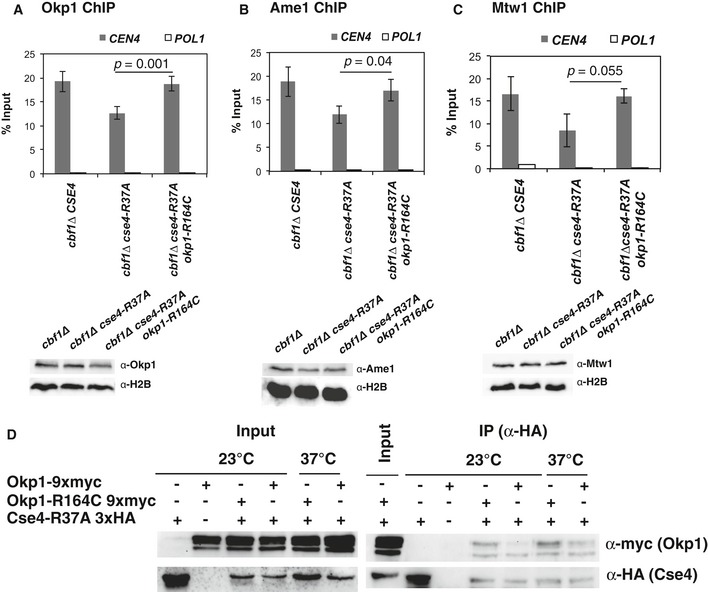
okp1‐R164C restored binding of Okp1 to centromeric sequences. ChIP of 9xmyc‐tagged Okp1 was performed in strains with the indicated genotypes that were grown at 37°C for 4 h before ChIP. Enrichment of CEN4 and POL1 (as a control) relative to input is given. Below, Western blot analysis of the amounts of 9xmyc‐Okp1 and histone H2B (loading control) in whole cell extracts.
okp1‐R164C restored binding of Ame1 to centromeric sequences. ChIP of 9xmyc‐tagged Ame1 was performed as in (A). Below, Western blot analysis of the amounts of 9xmyc‐Ame1 and H2B as in (A).
okp1‐R164C restored binding of Mtw1 to centromeric sequences. ChIP of 9xmyc‐Mtw1 and Western blots (right) are presented as in (A) and (B).
okp1‐R164C improved in vivo association of Okp1 with Cse4‐R37A. Co‐immunoprecipitation of Cse4‐R37A and Okp1 was carried out in cells carrying cse4‐R37A with (+) or without (−) 3xHA‐tag and OKP1 or okp1‐R164C with (+) or without (−) 9xmyc‐tag (AEY5040, AEY5972, AEY5973, AEY6589, AEY6592). Cells were grown at 23°C or were shifted to 37°C for 5 h prior to harvesting. Cse4‐R37A was immunoprecipitated using an α‐HA antibody, and the presence of Cse4‐R37A and Okp1 or Okp1‐R164C in the immunoprecipitate was tested by Western blotting with α‐HA and α‐myc antibody, respectively. Left, inputs (the two bands indicate Okp1 and a shorter degradation product); right, α‐HA immunoprecipitates.
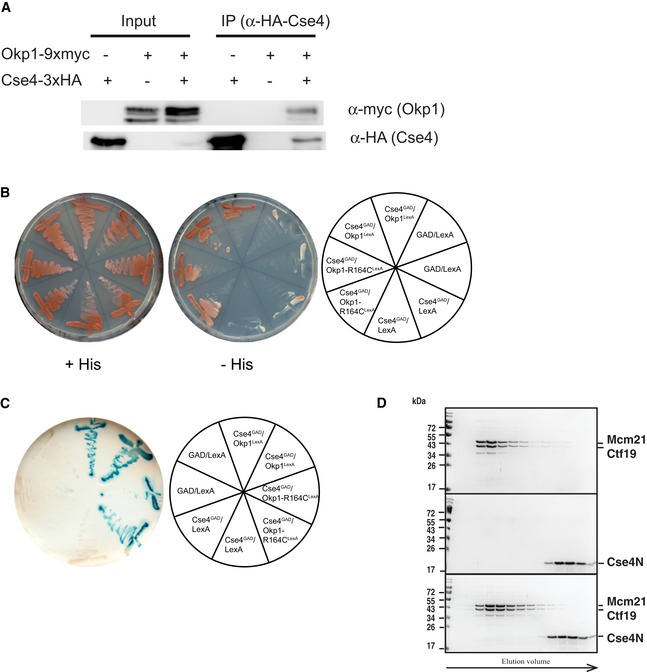
Co‐immunoprecipitation of Okp1 and Cse4. Co‐IP was carried out in cells carrying CSE4 with (+) or without (−) 3xHA‐tag and OKP1 with (+) or without (−) 9xmyc‐tag. Cells were grown at 30°C, and Cse4 was immunoprecipitated using an α‐HA antibody (AEY2781, AEY5972, AEY6578). The presence of Cse4 and Okp1 in the immunoprecipitate was tested by Western blotting with α‐HA and α‐myc antibody, respectively. Left: inputs of the indicated strains; right: α‐HA immunoprecipitates of the indicated strains.
Two‐hybrid analysis of the interaction between Cse4 (aa 11–139) and Okp1. Cells carrying the indicated constructs were streaked on minimal medium (+His) or on medium lacking histidine (−His) to test for activation of the two‐hybrid reporter.
Activity of the lacZ two‐hybrid reporter of strains carrying the indicated two‐hybrid constructs.
Absence of in vitro interaction between Ctf19/Mcm21 and Cse4N. Recombinant Ctf19/Mcm21 and Cse4N were co‐incubated prior to size‐exclusion chromatography (SEC) as for Okp1/Ame1 in Fig 4A. Coomassie Blue‐stained SDS–PAGE gels of SEC runs with Mcm21/Ctf19 (top), Cse4N (middle) and co‐incubation of Ctf19/Mcm21 with Cse4N (bottom) are shown.
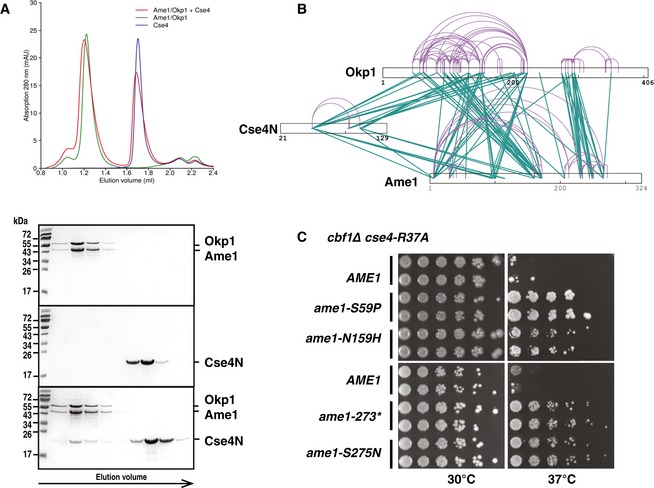
Cse4 (21–129, Cse4N) and Okp1/Ame1 were expressed and purified separately from bacteria and co‐incubated before separation by analytical gel filtration. Top, representative size‐exclusion chromatography (SEC) chromatogram with absorbance measurements at 280 nanometres (nm) is shown. Below, image of Coomassie Blue‐stained SDS–PAGE gels with fractions from principal SEC peaks of runs with Okp1/Ame1 alone (top), Cse4N alone (middle) and Okp1/Ame1 preincubated with Cse4N before SEC (bottom). Molecular weight standards are shown in kilodalton (kDa).
Intersubunit and self‐link map for the Cse4N/Okp1/Ame1 complex formed in vitro show direct contacts of Cse4N with both Okp1 and Ame1. Self‐links and intersubunit cross‐links are coloured in purple and green, respectively.
The ame1 mutations S59P, N159H, S275N and a C‐terminal truncation after aa 273 (ame1‐273*) suppressed the growth defect of cbf1∆ cse4‐R37A. cbf1∆ cse4‐R37A ame1∆ strains carrying the indicated plasmid‐borne ame1 alleles are shown as in Fig 2B.
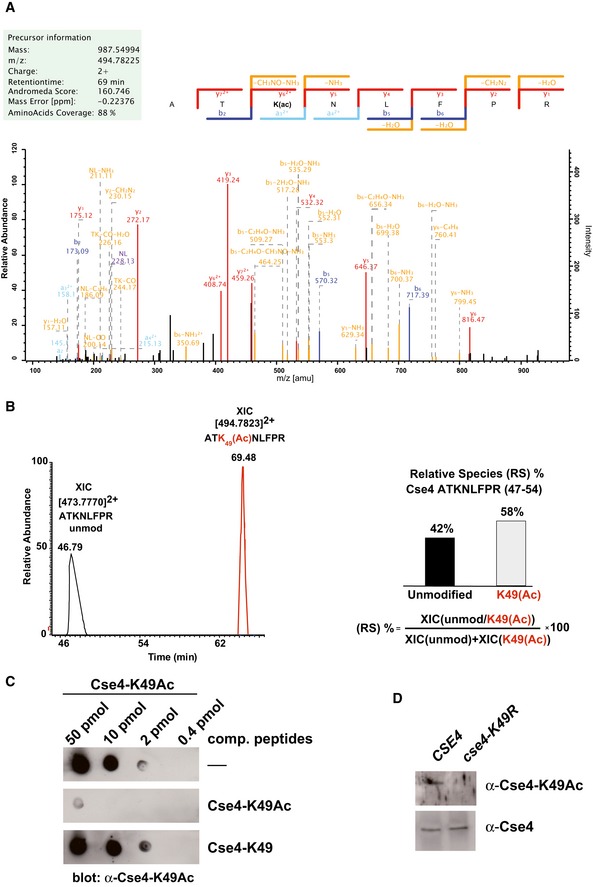
Collision‐induced dissociation analysis and full annotation of the parent ion (Cse4 47‐ATK(Ac)NLFPR‐54) with m/z = 494.78. Score and expected values as calculated by Mascot are reported, and fragments detected in the experiment are indicated in colour (blue for b ions and red for y ions).
Extract ion chromatograms (XIC) relative to each form of the peptide (left) were used to estimate relative abundance of the two species (right).
Specificity of the α‐Cse4‐K49Ac antibody. Immuno‐dot blot showing the specific reactivity of the antibody. The indicated amounts of Cse4‐K49Ac peptide were spotted on a nitrocellulose membrane. Antibodies were preincubated with the competing peptides indicated to the right of the blots.
Partially purified Cse4 (Waterborg, 2000; Samel et al, 2012) from WT (AEY2781) and cse4‐K49R (AEY2960) cells was probed with the α‐Cse4‐K49Ac antibody, as well as with α‐HA to detect 3xHA‐Cse4 as a loading control.
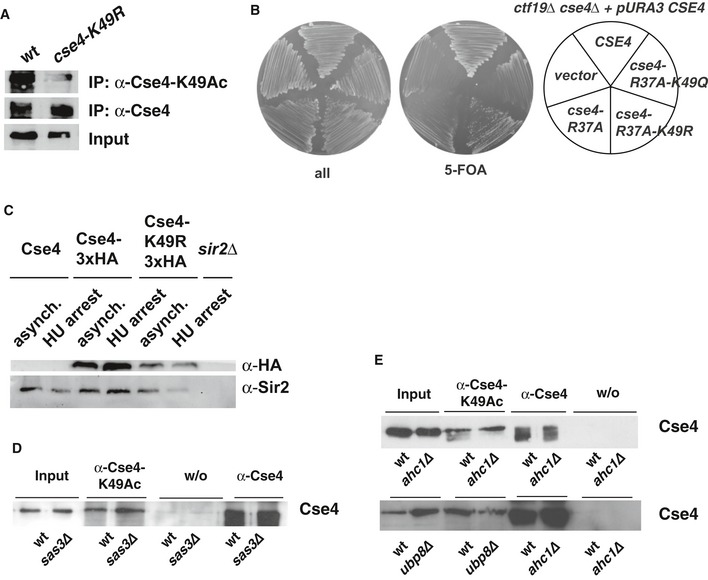
Cse4 is not precipitated with the α‐Cse4‐K49Ac in cells carrying cse4‐K49R. Cells carrying HA‐tagged Cse4 were used to immunoprecipitate Cse4 with an α‐Cse4‐K49Ac (top) and an α‐HA antibody (bottom).
The growth defect of cbf1∆ cse4‐R37A was not suppressed by additional cse4‐K49Q mutation.
Mutation of Cse4‐K49R did not affect Cse4 degradation. Western blot analysis of the amounts of Cse4‐3xHA and Sir2 (loading control) in whole cell extracts of asynchronous cultures or cells arrested with hydroxyurea (HU, 3 h arrest). Strains used were as follows: Cse4, AEY1; Cse4‐3xHA, AEY2781; Cse4‐K49R‐3xHA, AEY2960; sir2∆, AEY13. Extract of a sir2∆ strain was included to show specificity of the Sir2 antibody.
Acetylation of Cse4‐K49 is independent of the HAT Sas3. Immunoprecipitations (IP) were performed as in (A) using cells with sas3∆.
Ahc1 and Ubp8 are not required for Cse4‐K49Ac in vivo. IPs were performed as in (A) using ahc1∆ and ubp8∆ strains.
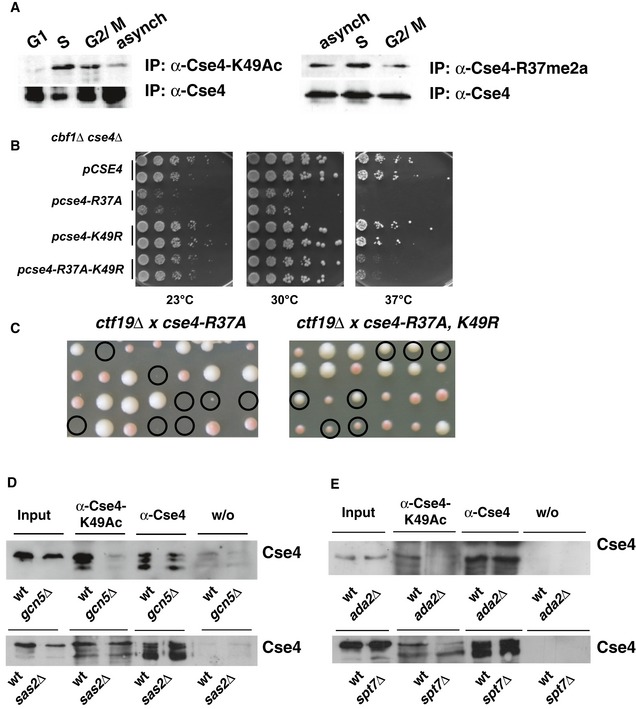
Cse4‐K49Ac and –R37Me are increased in S‐phase of the cell cycle. Cells carrying 3xHA‐tagged Cse4 were used to immunoprecipitate Cse4 with an α‐Cse4‐K49Ac (left, top) and an α‐HA antibody (left, bottom). To detect Cse4‐R37 methylation, 3xHA‐tagged Cse4 was immunoprecipitated with an α‐Cse4‐R37Me2a (right, top; Samel et al, 2012) and an α‐HA antibody (right, bottom). Western blots were probed for the presence of Cse4 with the α‐HA antibody. G1, treatment with α‐factor; S, arrest with hydroxyurea; G2/M, arrest in nocodazole; asynch, asynchronous culture.
Mimicking the unacetylated state of Cse4‐K49 (cse4‐K49R) partially suppressed the temperature sensitivity of cbf1∆ cse4‐R37A. cbf1∆ cse4∆ strains carrying the indicated cse4 alleles on a plasmid were spotted on full medium and grown as in Fig 2B.
cse4‐K49R suppressed the lethality of cse4‐R37A with ctf19∆. Tetrad dissection of a ctf19∆ strain crossed with cse4‐R37A (left) and cse4‐R37A K49R (right). Representation as in Fig 2D.
Acetylation of Cse4‐K49 was reduced in the absence of Gcn5 and the SAGA complex, but not in the absence of the HAT Sas2. Cse4 was precipitated as in (A) from strains wild‐type (wt) or gcn5∆ (top) or sas2∆ strains (bottom) carrying HA‐tagged Cse4. w/o, control immunoprecipitation without antibody.
Cse4‐K49Ac depended on the SAGA components Ada2 and Spt7. Cse4‐K49Ac was determined as in (A, left) with cells carrying ada2∆ or spt7∆.
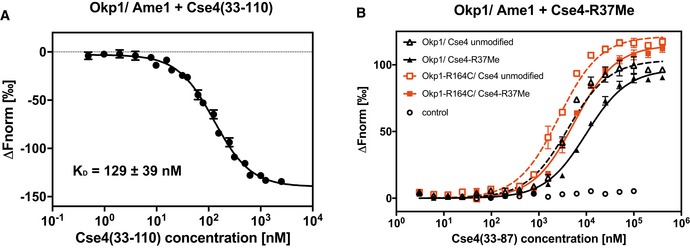
A synthetic 78‐aa Cse4 peptide (aa 33–110) binds to Okp1/Ame1 in vitro. Microscale thermophoresis (MST) measurement of binding of fluorescently labelled Cse4 (33–110) to Okp1/Ame1 is shown. Normalized fluorescence, from which the fluorescence of unbound Okp1/Ame1 was subtracted (∆F norm), is plotted against Cse4 (33–110) concentration (logarithmic scale). Mean values ± SD (n = 3) are shown.
Plot of MST measuring the competition of fluorescently labelled Cse4 (33–110) bound to Okp1/Ame1 (black; or Okp1‐R164C/Ame1, red) with unlabelled Cse4 (33–87) that is unmodified or monomethylated on R37 (R37Me). Normalized fluorescence from which the fluorescence of unbound Cse4 (33–110) was subtracted (∆F norm) is plotted against peptide concentration. Mean values ± standard deviation (n = 3) are given. Control, normalized fluorescence of Cse4 (33–110) alone at increasing concentrations.
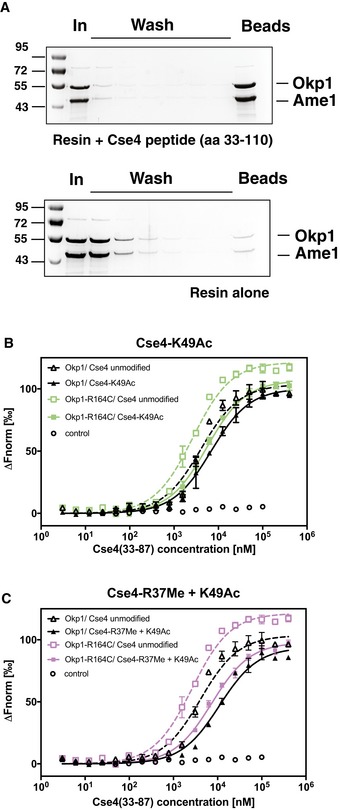
A synthetic 78‐aa Cse4 peptide (aa 33–110) binds to Okp1/Ame1 in vitro. Pull‐down of Okp1/Ame1 by Cse4 (33–110). The Cse4 peptide was coupled to sepharose beads, and recombinant Okp1/Ame1 was incubated with beads with (top) or without (bottom) coupled Cse4 peptide. Beads were washed, and remaining bound Okp1/Ame1 amounts were determined. Samples of input (In), washes and beads were separated on SDS–PAGE gels, which were stained with Coomassie Blue.
The binding of Okp1/Ame1 to Cse4N was inhibited by Cse4‐K49Ac, and Okp1‐R164C improved binding to Cse4N. Plot of MST measuring the competition of fluorescently labelled Cse4 (33–110) bound to Okp1/Ame1 (black; or Okp1‐R164C/Ame1, green) with unlabelled Cse4 (33–87) that is unmodified or acetylated on K49 (K49Ac). Data are represented as in Fig 6B.
Binding of Okp1/Ame1 to Cse4N was inhibited by simultaneous Cse4‐R37Me and ‐K49Ac. Representation as in (B), but using purple for Okp1‐R164C/Ame1.
References
-
- Angus‐Hill ML, Dutnall RN, Tafrov ST, Sternglanz R, Ramakrishnan V (1999) Crystal structure of the histone acetyltransferase Hpa2: a tetrameric member of the Gcn5‐related N‐acetyltransferase superfamily. J Mol Biol 294: 1311–1325 - PubMed
-
- Boeckmann L, Takahashi Y, Au WC, Mishra PK, Choy JS, Dawson AR, Szeto MY, Waybright TJ, Heger C, McAndrew C, Goldsmith PK, Veenstra TD, Baker RE, Basrai MA (2013) Phosphorylation of centromeric histone H3 variant regulates chromosome segregation in Saccharomyces cerevisiae . Mol Biol Cell 24: 2034–2044 - PMC - PubMed
-
- Boeke JD, LaCroute F, Fink GR (1984) A positive selection for mutants lacking orotidine‐5′‐phosphate decarboxylase activity in yeast: 5‐fluoro‐orotic acid resistance. Mol Gen Genet 197: 345–346 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
