

Evolution of drug resistance in an antifungal-naive chronic Candida lusitaniae infection
- PMID: 30389707
- PMCID: PMC6255150
- DOI: 10.1073/pnas.1807698115
Evolution of drug resistance in an antifungal-naive chronic Candida lusitaniae infection
Abstract
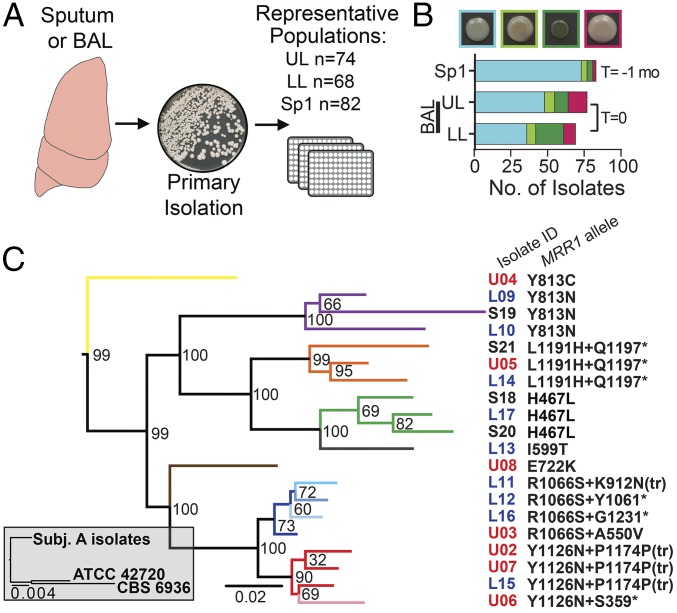
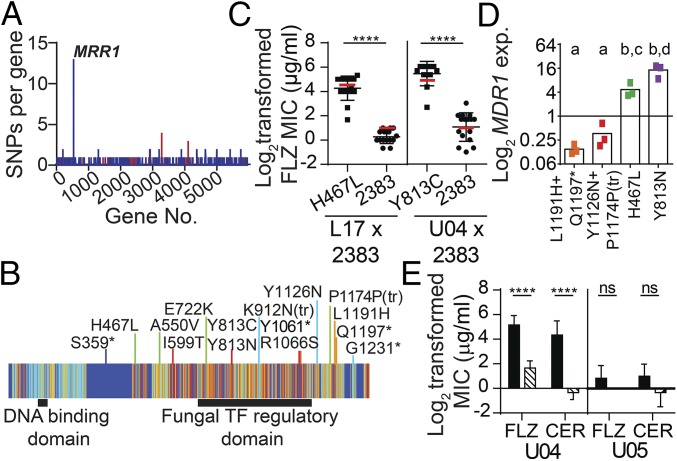
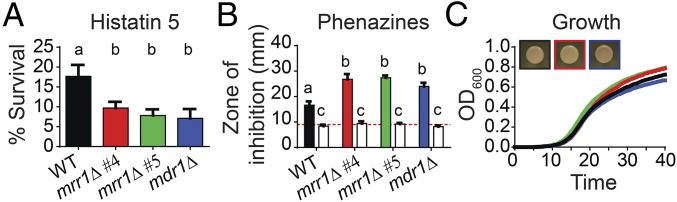
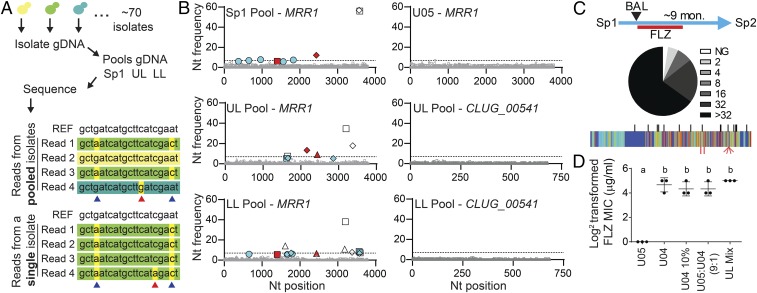
Management of the limited number of antimicrobials currently available requires the identification of infections that contain drug-resistant isolates and the discovery of factors that promote the evolution of drug resistance. Here, we report a single fungal infection in which we have identified numerous subpopulations that differ in their alleles of a single gene that impacts drug resistance. The diversity at this locus was markedly greater than the reported heterogeneity of alleles conferring antibiotic resistance in bacterial infections. Analysis of genomes from hundreds of Clavispora (Candida) lusitaniae isolates, through individual and pooled isolate sequencing, from a single individual with cystic fibrosis revealed at least 25 nonsynonymous mutations in MRR1, which encodes a transcription factor capable of inducing fluconazole (FLZ) resistance in Candida species. Isolates with high-activity Mrr1 variants were resistant to FLZ due to elevated expression of the MDR1-encoded efflux pump. We found that high Mrr1-regulated Mdr1 activity protected against host and bacterial factors, suggesting drug resistance can be selected for indirectly and perhaps explaining the Mrr1 heterogeneity in this individual who had no prior azole exposure. Regional analysis of C. lusitaniae populations from the upper and lower lobes of the right lung suggested intermingling of subpopulations throughout. Our retrospective characterization of sputum and lung populations by pooled sequencing found that alleles that confer FLZ resistance were a minority in each pool, possibly explaining why they were undetected before unsuccessful FLZ therapy. New susceptibility testing regimes may detect problematical drug-resistant subpopulations in heterogeneous single-species infections.
Keywords: Candida; drug resistance; evolution; fungi; heterogeneity.
Copyright © 2018 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kao CY, et al. Heteroresistance of Helicobacter pylori from the same patient prior to antibiotic treatment. Infect Genet Evol. 2014;23:196–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
