

Evolution and Structural Characteristics of Plant Voltage-Gated K+ Channels
- PMID: 30389753
- PMCID: PMC6354262
- DOI: 10.1105/tpc.18.00523
Evolution and Structural Characteristics of Plant Voltage-Gated K+ Channels
Abstract
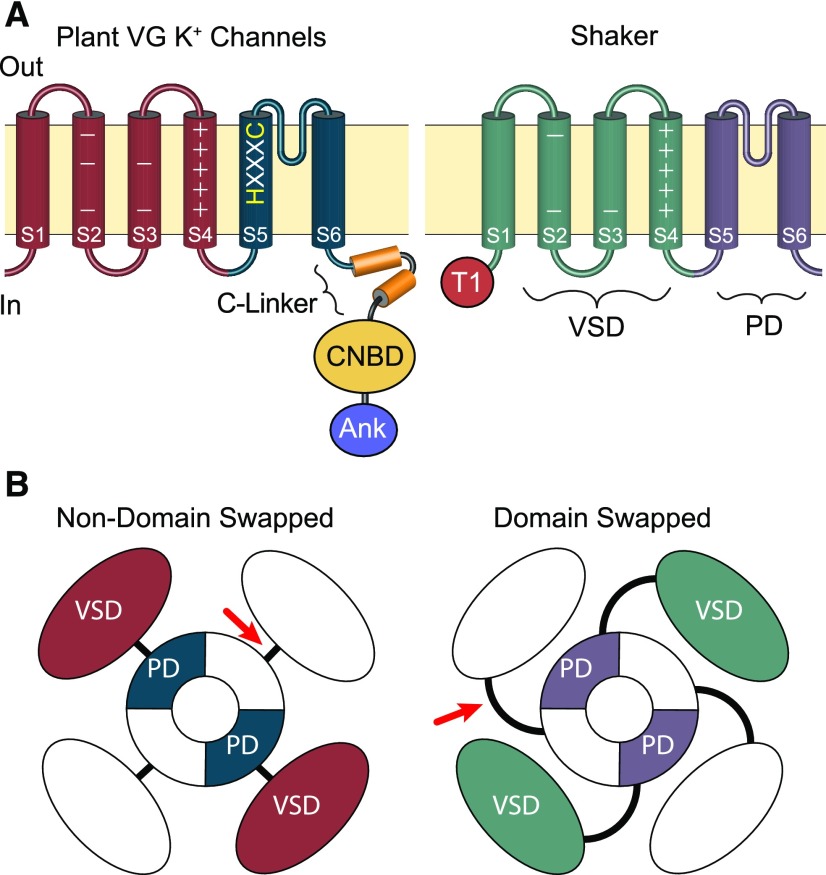
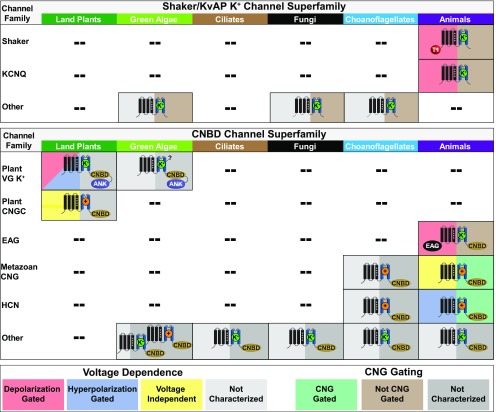
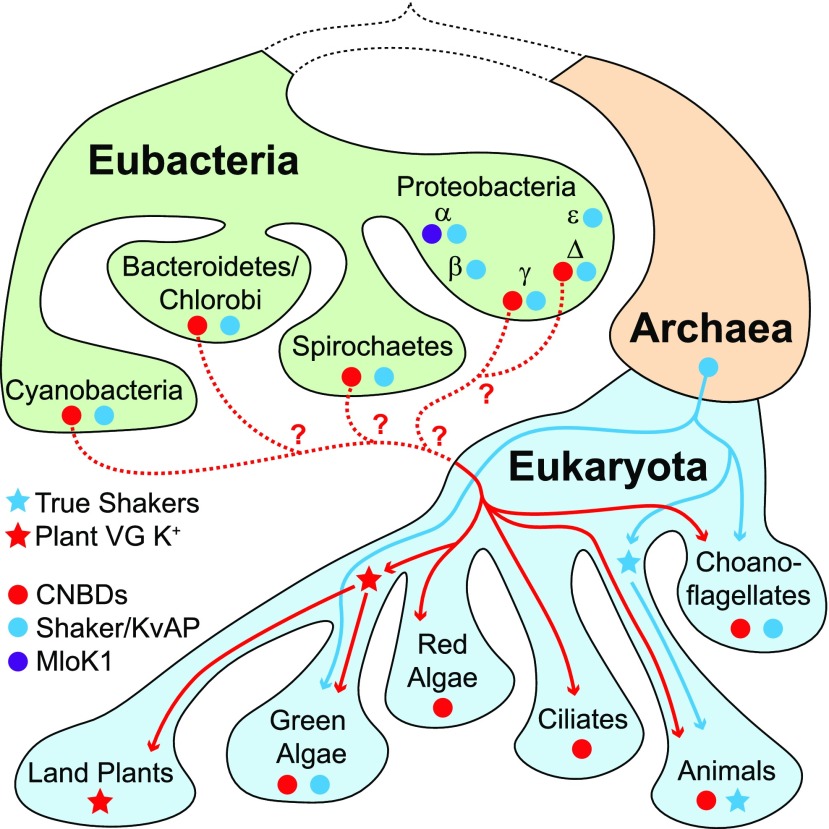
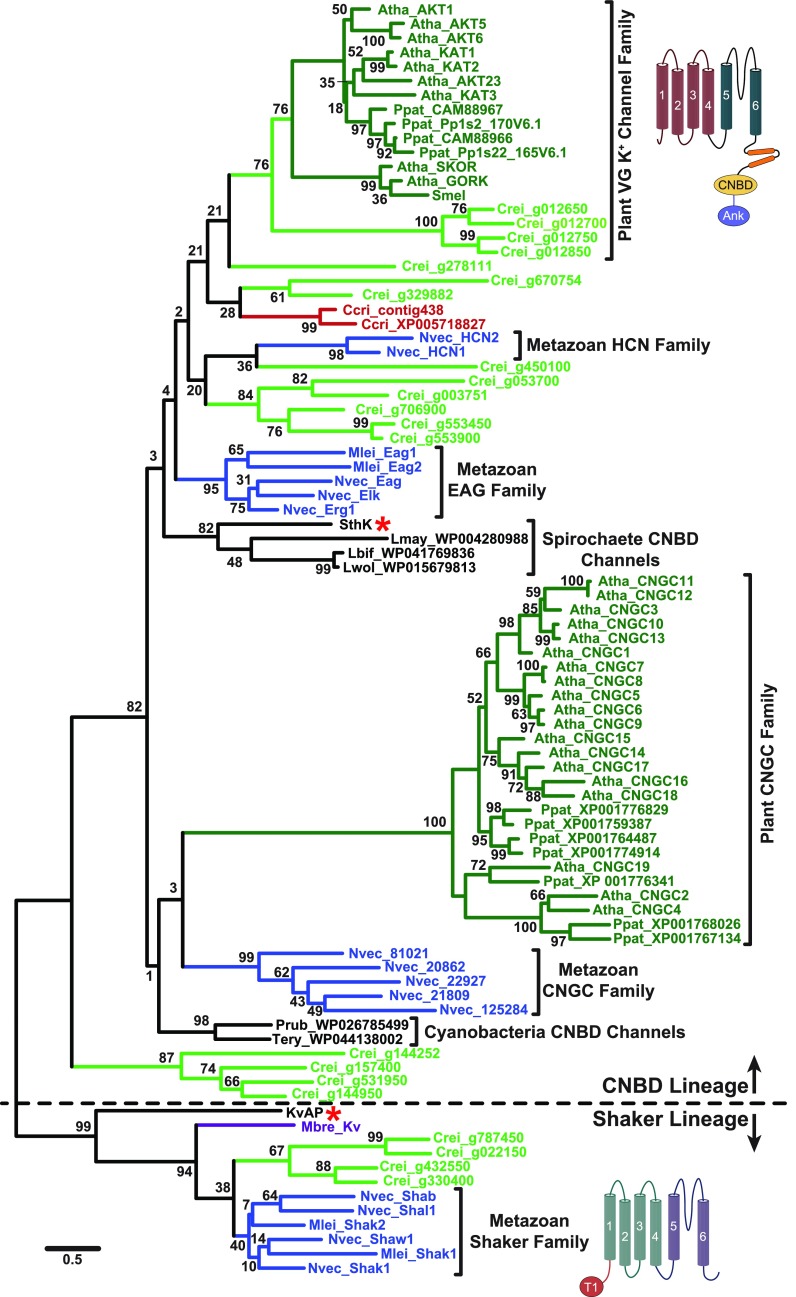
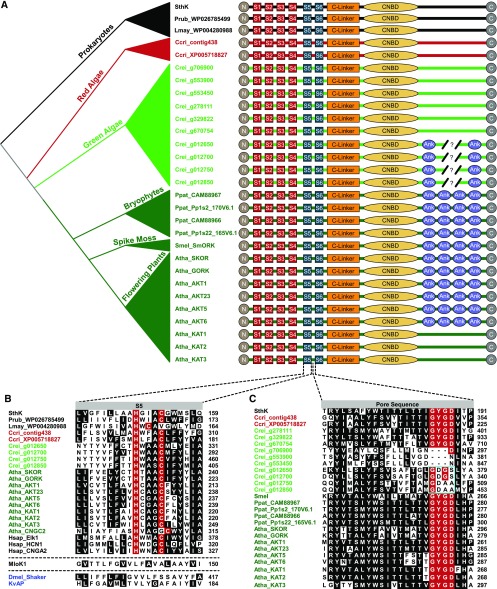
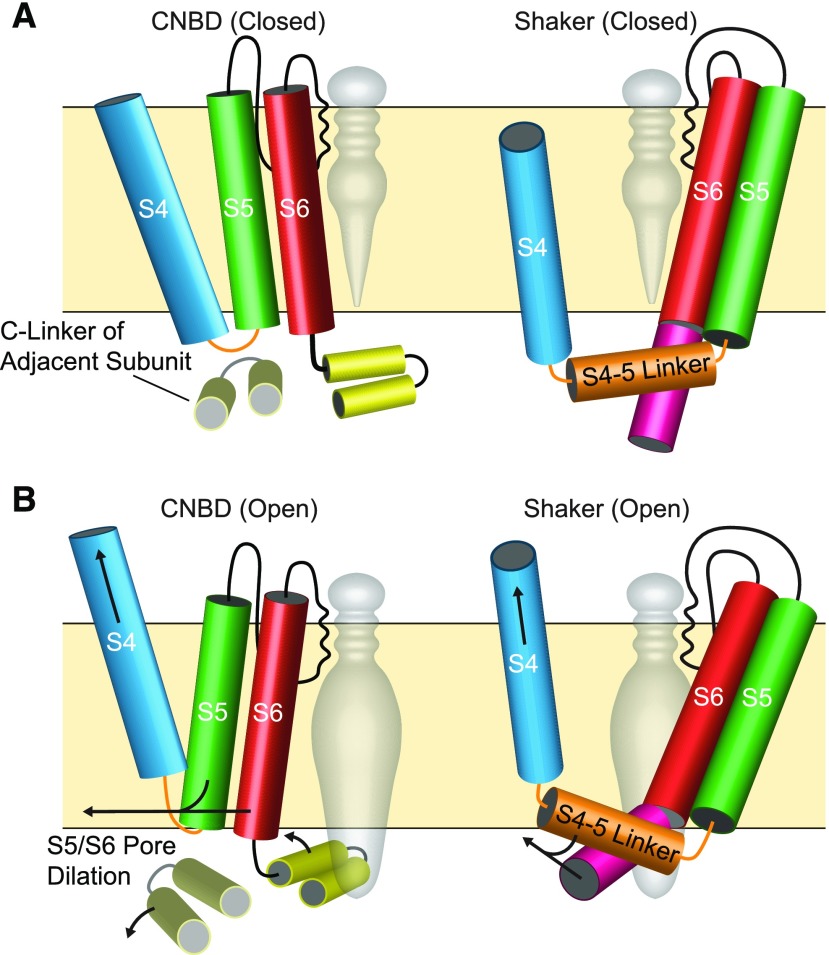
Plant voltage-gated K+ channels have been referred to as "plant Shakers" in reference to animal Shaker channels, the first K+ channels identified. Recent advances in our knowledge of K+ channel evolution and structure have significantly deepened the divide between these plant and animal K+ channels, suggesting that it is time to completely retire the "plant Shaker" designation. Evolutionary genomics reveals that plant voltage-gated K+ channels and metazoan Shakers derive from distinct prokaryotic ancestors. The plant channels belong to a lineage that includes cyclic nucleotide-gated channels and metazoan ether-à-go-go and hyperpolarization-activated, cyclic nucleotide-gated channels. We refer to this lineage as the CNBD channel superfamily, because all these channels share a cytoplasmic gating domain homologous to cyclic nucleotide binding domains. The first structures of CNBD superfamily channels reveal marked differences in coupling between the voltage sensor and ion-conducting pore relative to metazoan Shaker channels. Viewing plant voltage-gated K+ channel function through the lens of CNBD superfamily structures should lead to insights into how these channels are regulated.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
