

Embryos of an Antarctic zooplankton require anoxia for dormancy, are permeable to lipophilic chemicals, and reside in sediments containing PCBs
- PMID: 30390015
- PMCID: PMC6214904
- DOI: 10.1038/s41598-018-34689-w
Embryos of an Antarctic zooplankton require anoxia for dormancy, are permeable to lipophilic chemicals, and reside in sediments containing PCBs
Abstract
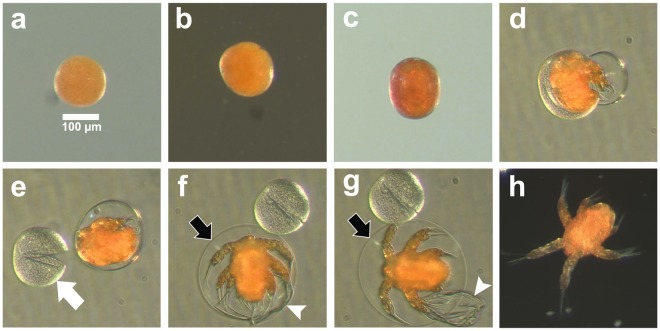
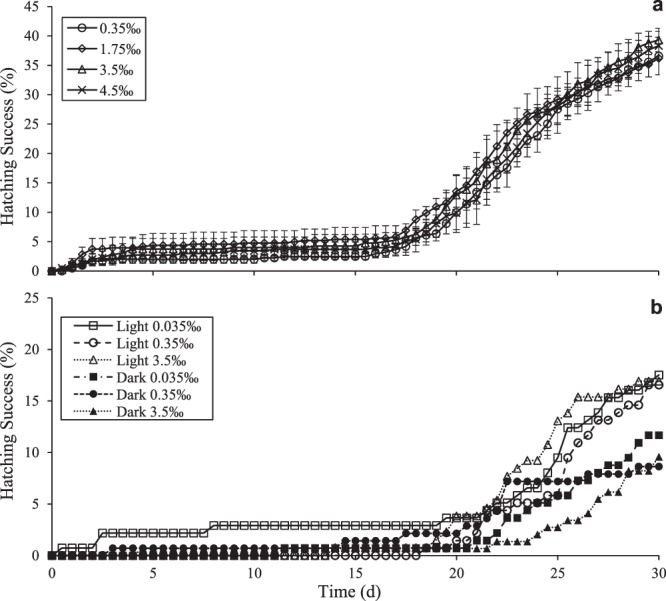
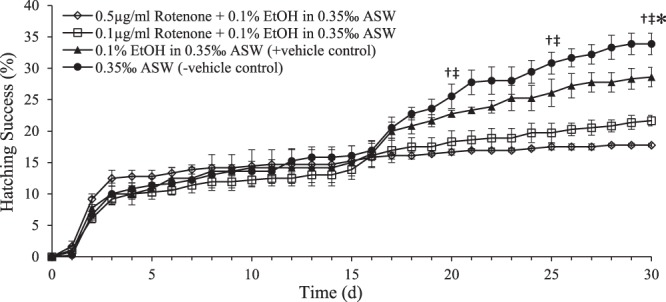
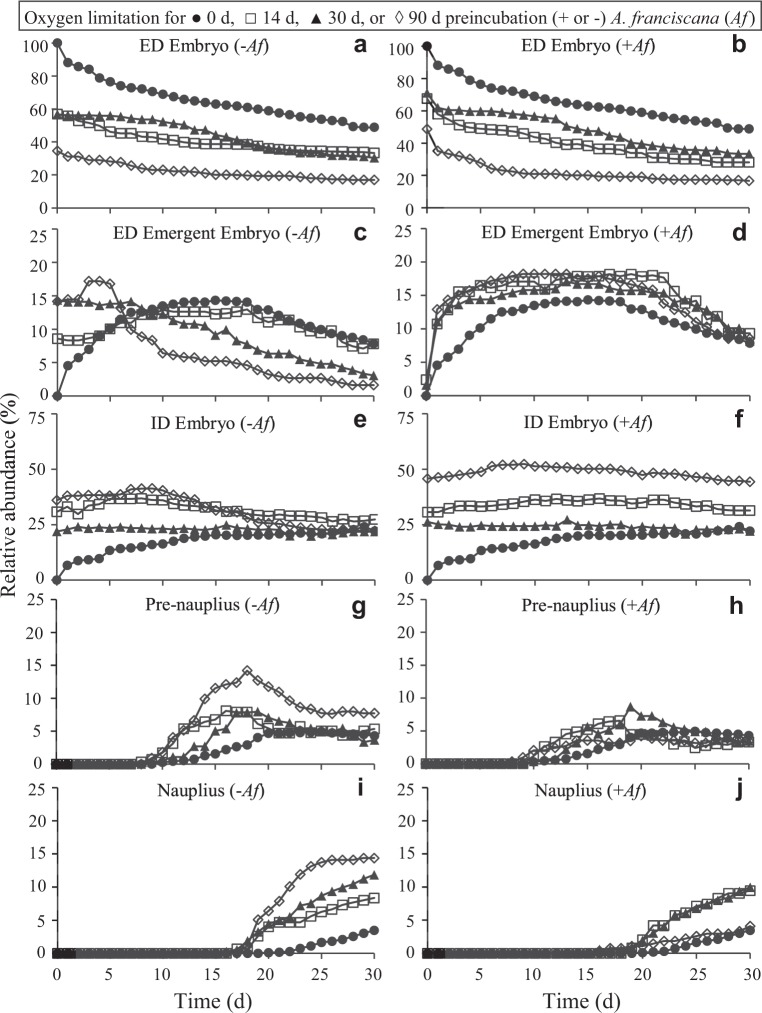
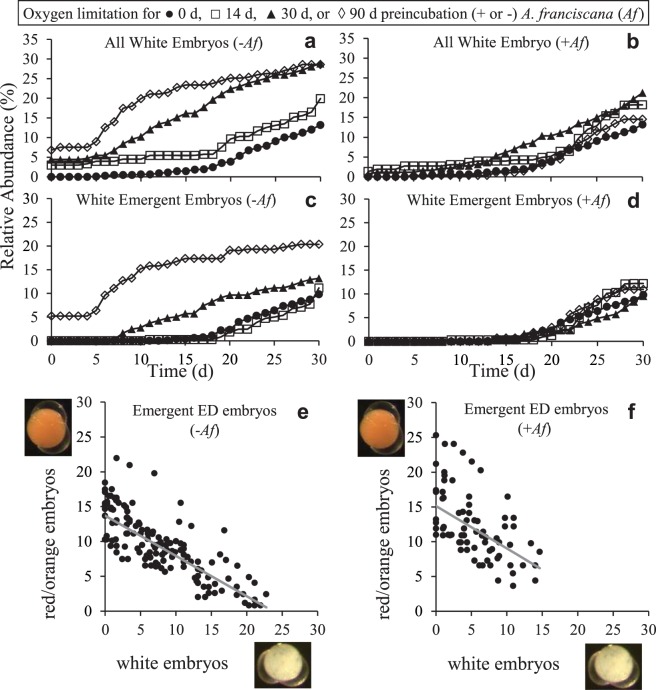
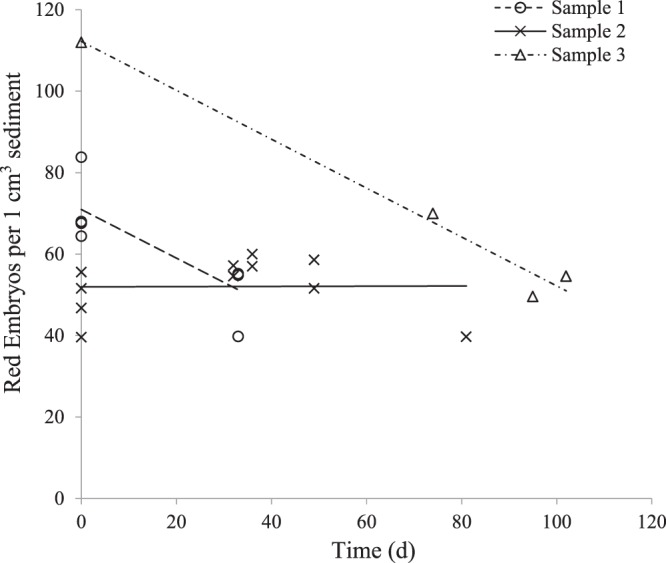
Zooplankton in Antarctic maritime lakes face challenges imposed by anthropogenic chemicals. Studies on temperate species suggest that lipophilic chemicals will accumulate in dormant embryos of Antarctic zooplankton and decrease hatching success, thereby threatening centuries of accumulated genetic diversity that would increase population resilience in the face of climate change. We evaluated the potential for lakes to act as sinks for legacy pollutants in the maritime Antarctic by testing sediments for polychlorinated biphenyls (PCBs) previously identified in soil, flora and fauna of lake catchments. Direct tests of embryo permeability to chemicals are confounded by potential adhesion of chemicals to the embryo surface and limited biomass available. Therefore, in order to assess the potential for lipophilic chemicals to penetrate and passively accumulate in dormant embryos of Antarctic lacustrine zooplankton, we evaluated the effect of anoxia on post-diapause development in the calanoid copepod, Boeckella poppei, and then used chemical anoxia induced by rotenone as a reporter for permeability of these embryos to moderately lipophilic chemicals. The data presented demonstrate that embryos of B. poppei from Antarctic lake sediments will passively accumulate moderately lipophilic chemicals while lying dormant in anoxic sediments. Implications for legacy POPs in sediments of Antarctic maritime lakes are discussed.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Reversible intracellular acidification and depletion of NTPs provide a potential physiological origin for centuries of dormancy in an Antarctic freshwater copepod.Sci Rep. 2023 Aug 15;13(1):13243. doi: 10.1038/s41598-023-40180-y. Sci Rep. 2023. PMID: 37582969 Free PMC article.
-
The ultrastructure of resurrection: Post-diapause development in an Antarctic freshwater copepod.J Struct Biol. 2021 Mar;213(1):107705. doi: 10.1016/j.jsb.2021.107705. Epub 2021 Feb 9. J Struct Biol. 2021. PMID: 33577904
-
Assessment of the cyst wall and surface microbiota in dormant embryos of the Antarctic calanoid copepod, Boeckella poppei.Environ Microbiol Rep. 2024 Dec;16(6):e70035. doi: 10.1111/1758-2229.70035. Environ Microbiol Rep. 2024. PMID: 39603712 Free PMC article.
-
Persistent organic pollutants in typical lake ecosystems.Ecotoxicol Environ Saf. 2019 Sep 30;180:668-678. doi: 10.1016/j.ecoenv.2019.05.060. Epub 2019 May 27. Ecotoxicol Environ Saf. 2019. PMID: 31146153 Review.
-
Polychlorinated biphenyls (PCBs) in bottom sediments: identification of sources.Chemosphere. 2014 Sep;111:151-6. doi: 10.1016/j.chemosphere.2014.03.025. Epub 2014 Apr 20. Chemosphere. 2014. PMID: 24997912 Review.
Cited by
-
Reversible intracellular acidification and depletion of NTPs provide a potential physiological origin for centuries of dormancy in an Antarctic freshwater copepod.Sci Rep. 2023 Aug 15;13(1):13243. doi: 10.1038/s41598-023-40180-y. Sci Rep. 2023. PMID: 37582969 Free PMC article.
References
-
- Marcus NH, Boero F. Minireview: The importance of benthic-pelagic coupling and the forgotten role of life cycles in coastal aquatic systems. Limnol. Oceanogr. 1998;43:763–768. doi: 10.4319/lo.1998.43.5.0763. - DOI
-
- Brendonck L, De Meester L. Egg banks in freshwater zooplankton: evolutionary and ecological archives in the sediment. Hydrobiologia. 2003;491:65–84. doi: 10.1023/A:1024454905119. - DOI
-
- Pociecha A, Dumont HJ. Life cycle of Boeckella poppei Mrazek and Branchinecta gaini Daday (King George Island, South Shetlands) Polar Biol. 2008;31:245–248. doi: 10.1007/s00300-007-0360-5. - DOI
-
- Jiang XD, et al. Abundance and age of viable resting eggs of the calanoid copepod Boeckella poppei Mrazek in sediments: evidence of egg banks in two Antarctic maritime lakes. Polar Biol. 2012;35:1525–1531. doi: 10.1007/s00300-012-1192-5. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
