

What Is the Relationship between Dopamine and Effort?
- PMID: 30391016
- PMCID: PMC6352317
- DOI: 10.1016/j.tins.2018.10.001
What Is the Relationship between Dopamine and Effort?
Abstract
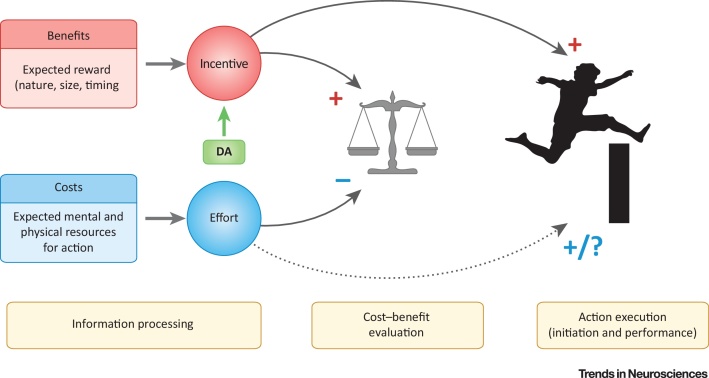
The trade-off between reward and effort is at the heart of most behavioral theories, from ecology to economics. Compared to reward, however, effort remains poorly understood, both at the behavioral and neurophysiological levels. This is important because unwillingness to overcome effort to gain reward is a common feature of many neuropsychiatric and neurological disorders. A recent surge in interest in the neurobiological basis of effort has led to seemingly conflicting results regarding the role of dopamine. We argue here that, upon closer examination, there is actually striking consensus across studies: dopamine primarily codes for future reward but is less sensitive to anticipated effort cost. This strong association between dopamine and the incentive effects of rewards places dopamine in a key position to promote reward-directed action.
Keywords: cost–benefit decision making; electrophysiology; midbrain; motivation; striatum; voltammetry.
Copyright © 2018 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Similar articles
-
Dopamine-Dependent Loss Aversion during Effort-Based Decision-Making.J Neurosci. 2020 Jan 15;40(3):661-670. doi: 10.1523/JNEUROSCI.1760-19.2019. Epub 2019 Nov 14. J Neurosci. 2020. PMID: 31727795 Free PMC article.
-
Blockade of dopamine D₁ receptors, but not D₂ receptors, decreases motivation in a novel effort-discounting paradigm in common marmosets.Behav Neurosci. 2018 Dec;132(6):526-535. doi: 10.1037/bne0000273. Epub 2018 Oct 8. Behav Neurosci. 2018. PMID: 30299146
-
Activational and effort-related aspects of motivation: neural mechanisms and implications for psychopathology.Brain. 2016 May;139(Pt 5):1325-47. doi: 10.1093/brain/aww050. Brain. 2016. PMID: 27189581 Free PMC article. Review.
-
The Neurobiology of Activational Aspects of Motivation: Exertion of Effort, Effort-Based Decision Making, and the Role of Dopamine.Annu Rev Psychol. 2024 Jan 18;75:1-32. doi: 10.1146/annurev-psych-020223-012208. Epub 2023 Oct 3. Annu Rev Psychol. 2024. PMID: 37788571 Review.
-
Striatal dopamine D2 receptors regulate effort but not value-based decision making and alter the dopaminergic encoding of cost.Neuropsychopharmacology. 2018 Oct;43(11):2180-2189. doi: 10.1038/s41386-018-0159-9. Epub 2018 Jul 20. Neuropsychopharmacology. 2018. PMID: 30082890 Free PMC article.
Cited by
-
Cocaine-induced increases in motivation require 2-arachidonoylglycerol mobilization and CB1 receptor activation in the ventral tegmental area.Neuropharmacology. 2021 Aug 1;193:108625. doi: 10.1016/j.neuropharm.2021.108625. Epub 2021 May 28. Neuropharmacology. 2021. PMID: 34058192 Free PMC article.
-
EngAge - A metacognitive intervention to supplement working memory training: A feasibility study in older adults.Aging Brain. 2023 Jul 7;4:100083. doi: 10.1016/j.nbas.2023.100083. eCollection 2023. Aging Brain. 2023. PMID: 38098966 Free PMC article.
-
Dopamine-Dependent Loss Aversion during Effort-Based Decision-Making.J Neurosci. 2020 Jan 15;40(3):661-670. doi: 10.1523/JNEUROSCI.1760-19.2019. Epub 2019 Nov 14. J Neurosci. 2020. PMID: 31727795 Free PMC article.
-
Neural signals implicated in the processing of appetitive and aversive events in social and non-social contexts.Front Syst Neurosci. 2022 Aug 3;16:926388. doi: 10.3389/fnsys.2022.926388. eCollection 2022. Front Syst Neurosci. 2022. PMID: 35993086 Free PMC article. Review.
-
Aversive motivation and cognitive control.Neurosci Biobehav Rev. 2022 Feb;133:104493. doi: 10.1016/j.neubiorev.2021.12.016. Epub 2021 Dec 12. Neurosci Biobehav Rev. 2022. PMID: 34910931 Free PMC article. Review.
References
-
- Berridge K.C. The debate over dopamine’s role in reward: the case for incentive salience. Psychopharmacology (Berl.) 2007;191:391–431. - PubMed
-
- Chong T.T., Husain M. The role of dopamine in the pathophysiology and treatment of apathy. Prog. Brain Res. 2016;229:389–426. - PubMed
-
- Robbins T.W., Everitt B.J. Functions of dopamine in the dorsal and ventral striatum. Semin. Neurosci. 1992;4:119–127.
-
- Phillips P.E. Calculating utility: preclinical evidence for cost–benefit analysis by mesolimbic dopamine. Psychopharmacology (Berl.) 2007;191:483–495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
