

Scale-Invariant Visual Capabilities Explained by Topographic Representations of Luminance and Texture in Primate V1
- PMID: 30392796
- PMCID: PMC6345167
- DOI: 10.1016/j.neuron.2018.10.020
Scale-Invariant Visual Capabilities Explained by Topographic Representations of Luminance and Texture in Primate V1
Erratum in
-
Scale-Invariant Visual Capabilities Explained by Topographic Representations of Luminance and Texture in Primate V1.Neuron. 2018 Dec 19;100(6):1533. doi: 10.1016/j.neuron.2018.12.011. Neuron. 2018. PMID: 30571944 Free PMC article. No abstract available.
Abstract
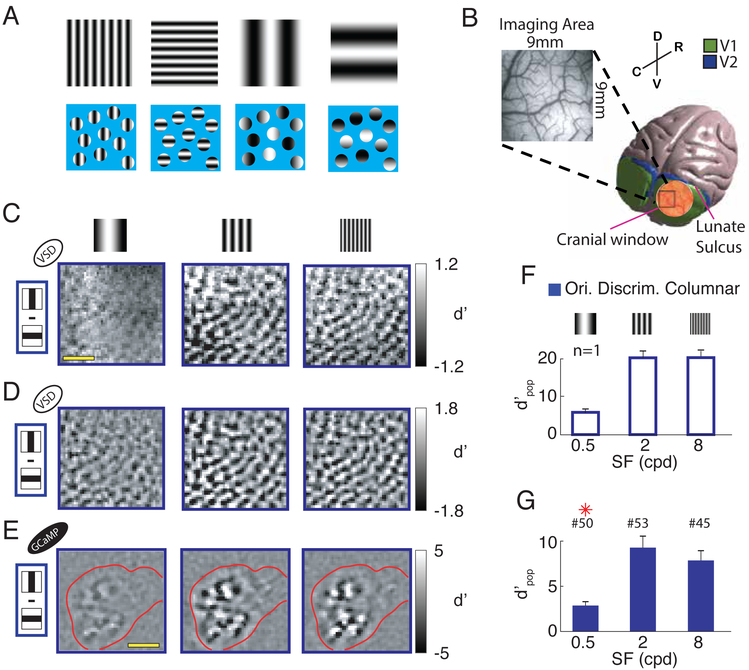
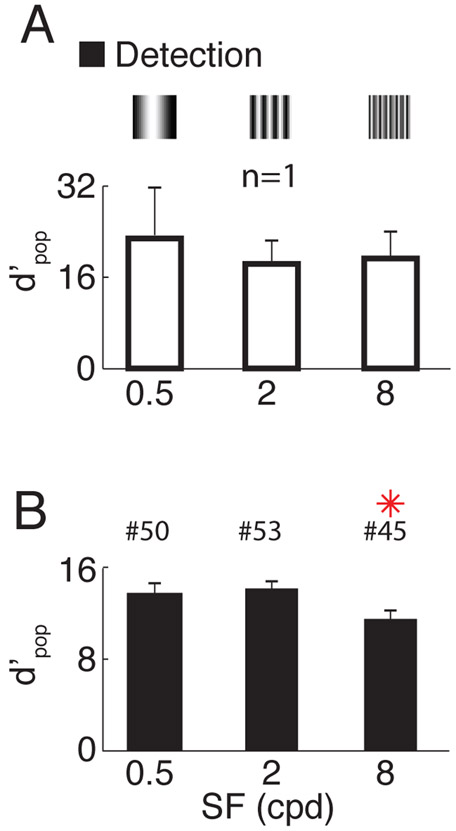
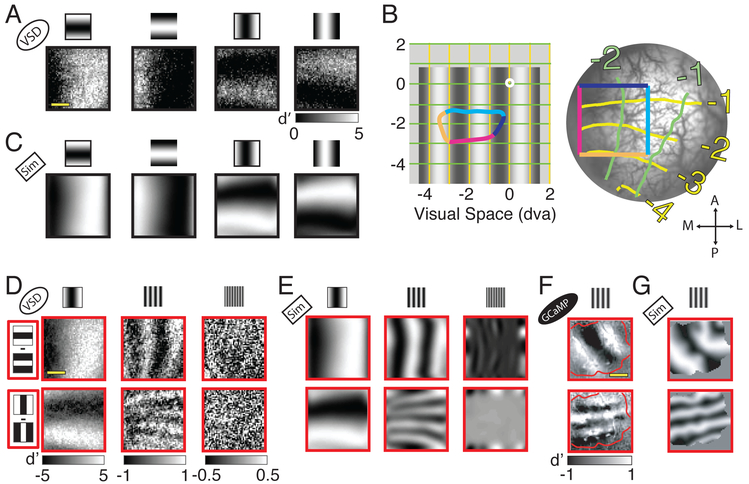
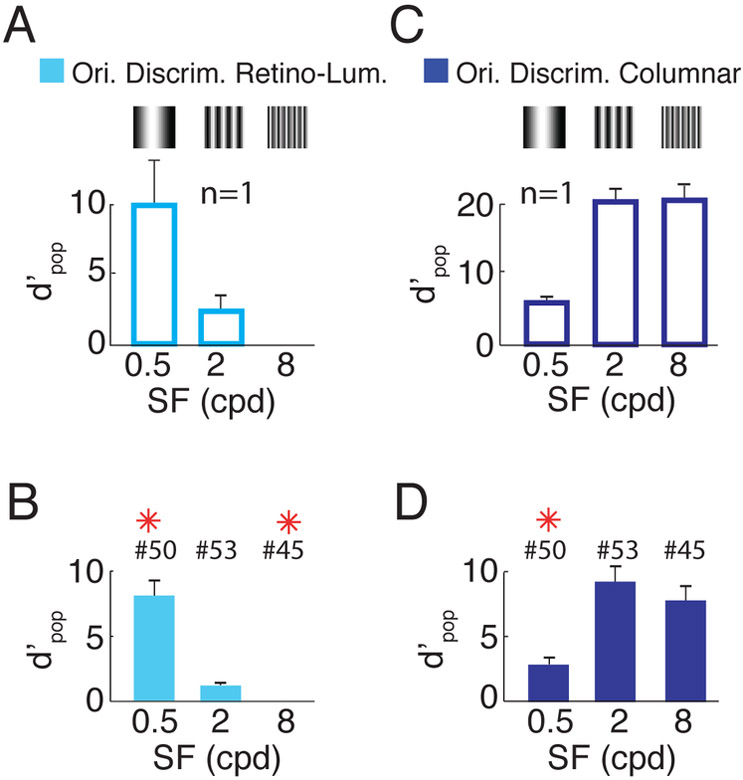
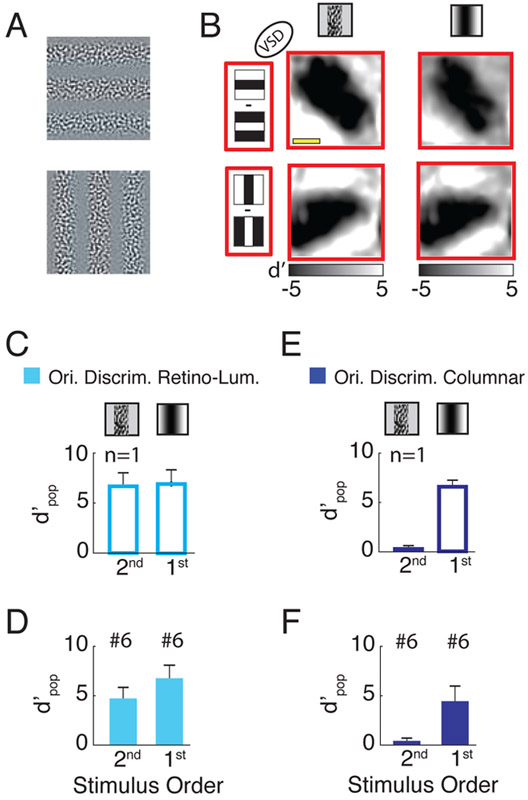
Humans have remarkable scale-invariant visual capabilities. For example, our orientation discrimination sensitivity is largely constant over more than two orders of magnitude of variations in stimulus spatial frequency (SF). Orientation-selective V1 neurons are likely to contribute to orientation discrimination. However, because at any V1 location neurons have a limited range of receptive field (RF) sizes, we predict that at low SFs V1 neurons will carry little orientation information. If this were the case, what could account for the high behavioral sensitivity at low SFs? Using optical imaging in behaving macaques, we show that, as predicted, V1 orientation-tuned responses drop rapidly with decreasing SF. However, we reveal a surprising coarse-scale signal that corresponds to the projection of the luminance layout of low-SF stimuli to V1's retinotopic map. This homeomorphic and distributed representation, which carries high-quality orientation information, is likely to contribute to our striking scale-invariant visual capabilities.
Keywords: genetically encoded cacium indicators; multi-scale representation; neural population code; optical imaging; orientation discrimination; orientation map; primary visual cortex; retinotopic map; visual perception; voltage sensitive dye.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Population encoding of spatial frequency, orientation, and color in macaque V1.J Neurophysiol. 1994 Nov;72(5):2151-66. doi: 10.1152/jn.1994.72.5.2151. J Neurophysiol. 1994. PMID: 7884450
-
Orientation-cue invariant population responses to contrast-modulated and phase-reversed contour stimuli in macaque V1 and V2.PLoS One. 2014 Sep 4;9(9):e106753. doi: 10.1371/journal.pone.0106753. eCollection 2014. PLoS One. 2014. PMID: 25188576 Free PMC article.
-
Cue-invariant detection of centre-surround discontinuity by V1 neurons in awake macaque monkey.J Physiol. 2007 Sep 1;583(Pt 2):581-92. doi: 10.1113/jphysiol.2007.130294. Epub 2007 Jun 28. J Physiol. 2007. PMID: 17599965 Free PMC article.
-
Functional cell classes and functional architecture in the early visual system of a highly visual rodent.Prog Brain Res. 2005;149:127-45. doi: 10.1016/S0079-6123(05)49010-X. Prog Brain Res. 2005. PMID: 16226581 Review.
-
Flexible information representation to stabilize sensory perception despite minor external input variations.Neurosci Res. 2023 Oct;195:1-8. doi: 10.1016/j.neures.2023.05.002. Epub 2023 May 25. Neurosci Res. 2023. PMID: 37236268 Review.
Cited by
-
Neural correlates of perceptual similarity masking in primate V1.Elife. 2024 Apr 9;12:RP89570. doi: 10.7554/eLife.89570. Elife. 2024. PMID: 38592269 Free PMC article.
-
Relationships between the degrees of freedom in the affine Gaussian derivative model for visual receptive fields and 2-D affine image transformations with application to covariance properties of simple cells in the primary visual cortex.Biol Cybern. 2025 Jun 18;119(2-3):15. doi: 10.1007/s00422-025-01014-4. Biol Cybern. 2025. PMID: 40531345 Free PMC article.
-
Uniform spatial pooling explains topographic organization and deviation from receptive-field scale invariance in primate V1.Nat Commun. 2020 Dec 15;11(1):6390. doi: 10.1038/s41467-020-19954-9. Nat Commun. 2020. PMID: 33319775 Free PMC article.
-
Mesoscopic calcium imaging in a head-unrestrained male non-human primate using a lensless microscope.Nat Commun. 2024 Feb 10;15(1):1271. doi: 10.1038/s41467-024-45417-6. Nat Commun. 2024. PMID: 38341403 Free PMC article.
-
Coding strategy for surface luminance switches in the primary visual cortex of the awake monkey.Nat Commun. 2022 Jan 12;13(1):286. doi: 10.1038/s41467-021-27892-3. Nat Commun. 2022. PMID: 35022404 Free PMC article.
References
-
- Adelson E, and Movshon J (1982). Phenomenal coherence of moving visual patterns. Nature 300, 523–525. - PubMed
-
- Adelson EH, and Bergen JR (1985). Spatiotemporal energy models for the perception of motion. J. Opt. Soc. Am. A 2, 284–299. - PubMed
-
- Angelucci A, Levitt JB, and Lund JS (2002). Anatomical origins of the classical receptive field of single neurons in macaque visual cortical area V1. Prog. Brain Res 136, 373–388. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
