

Inositol phosphate kinases: Expanding the biological significance of the universal core of the protein kinase fold
- PMID: 30392847
- PMCID: PMC9364425
- DOI: 10.1016/j.jbior.2018.10.006
Inositol phosphate kinases: Expanding the biological significance of the universal core of the protein kinase fold
Abstract
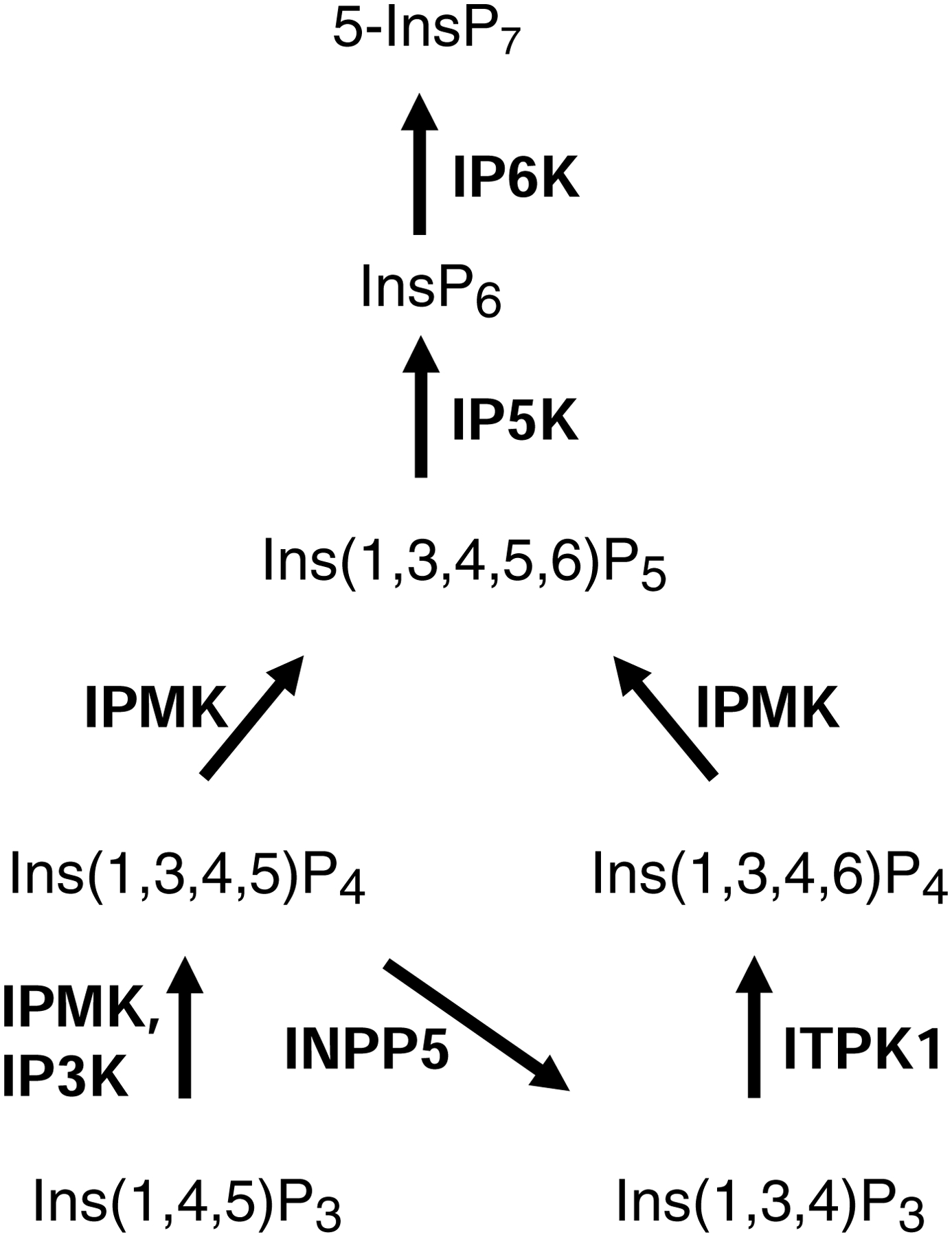
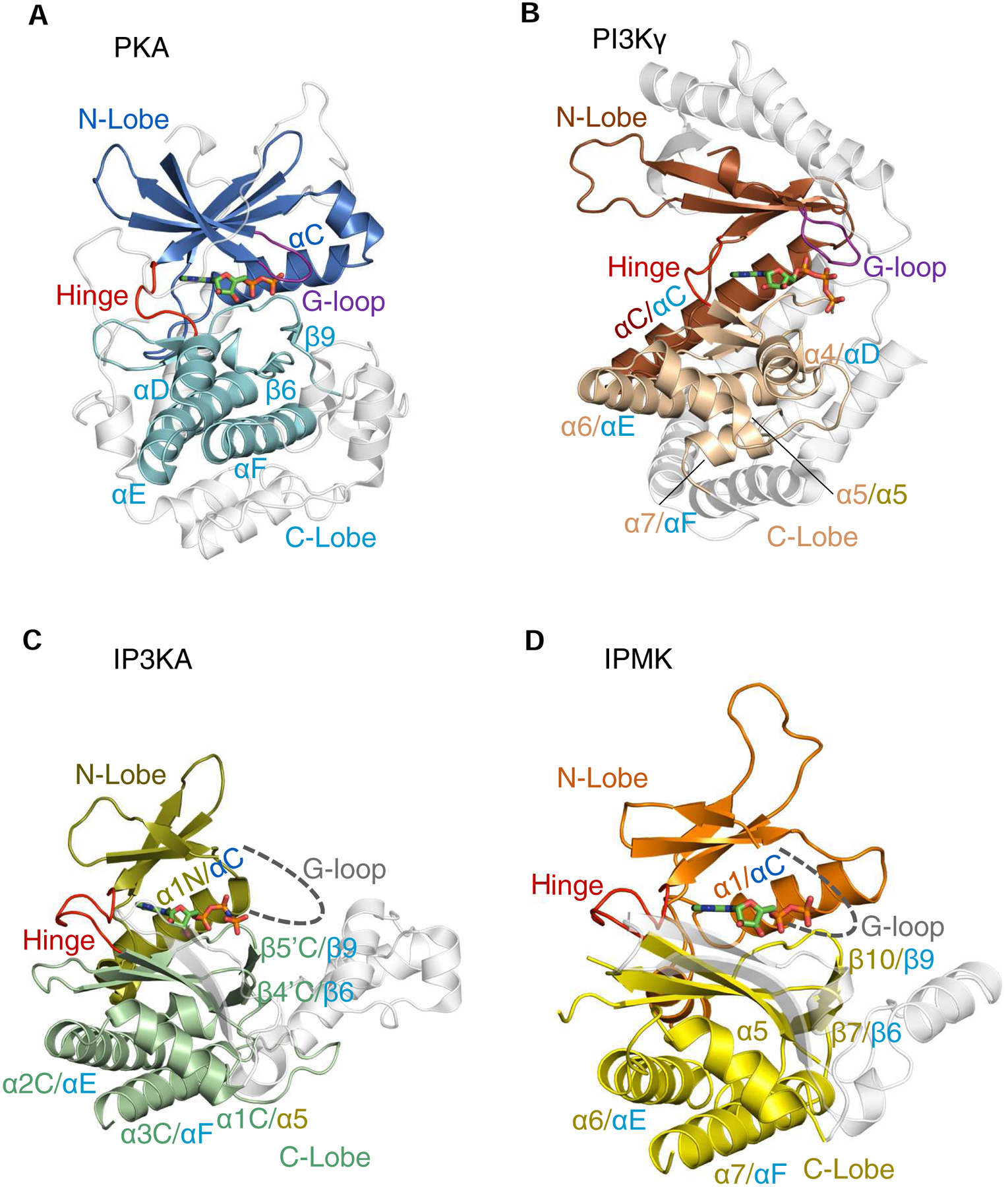
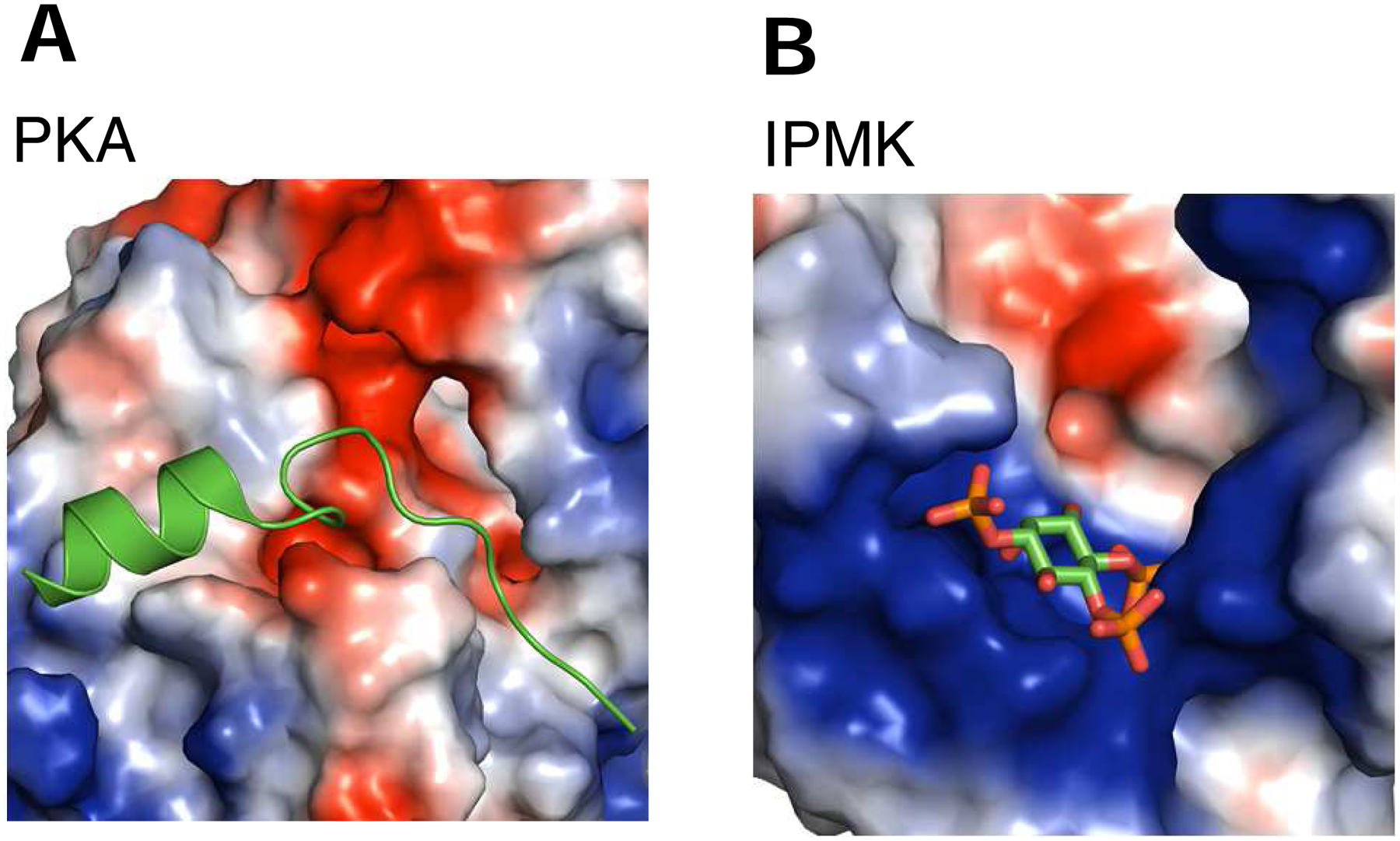
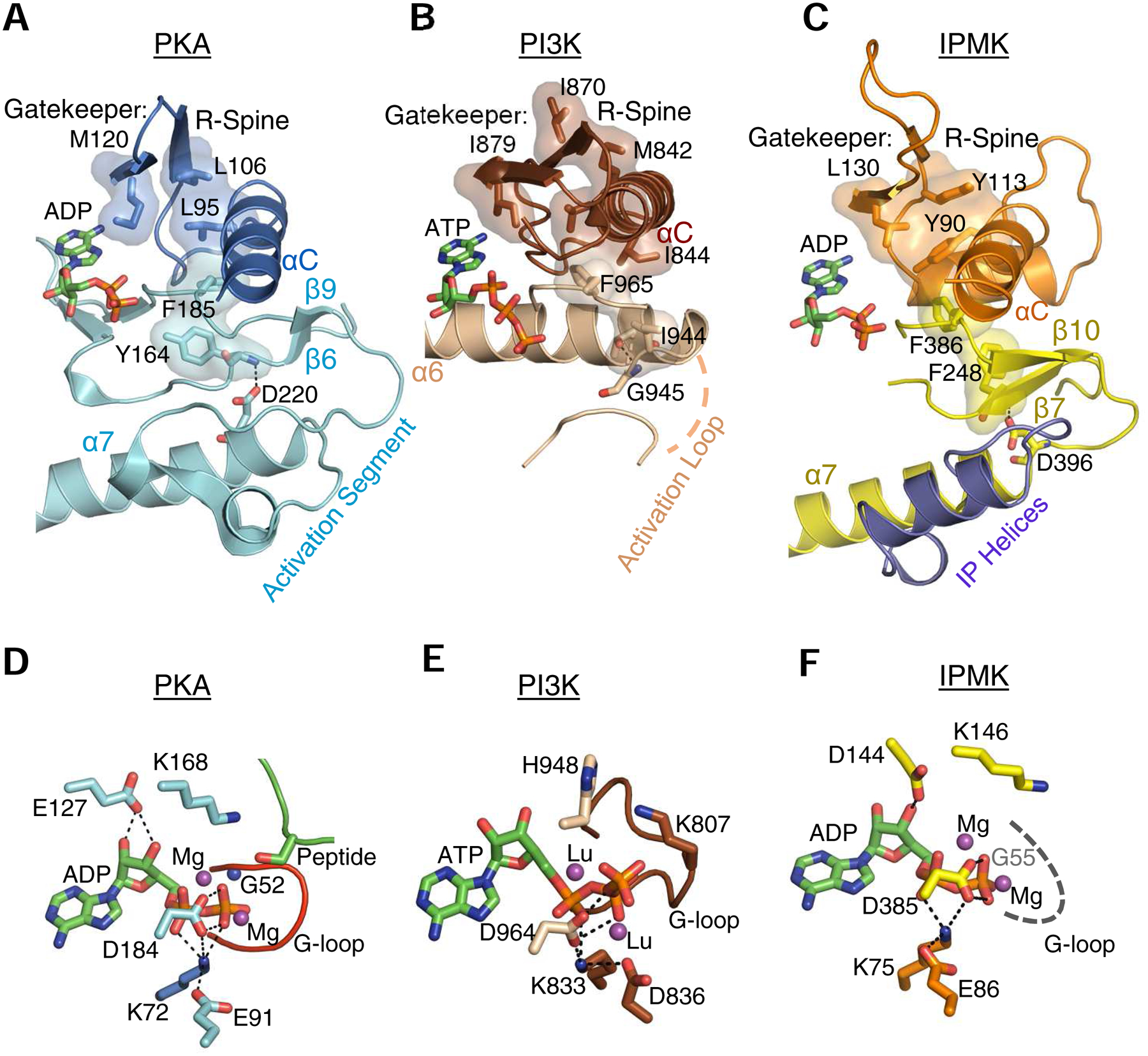
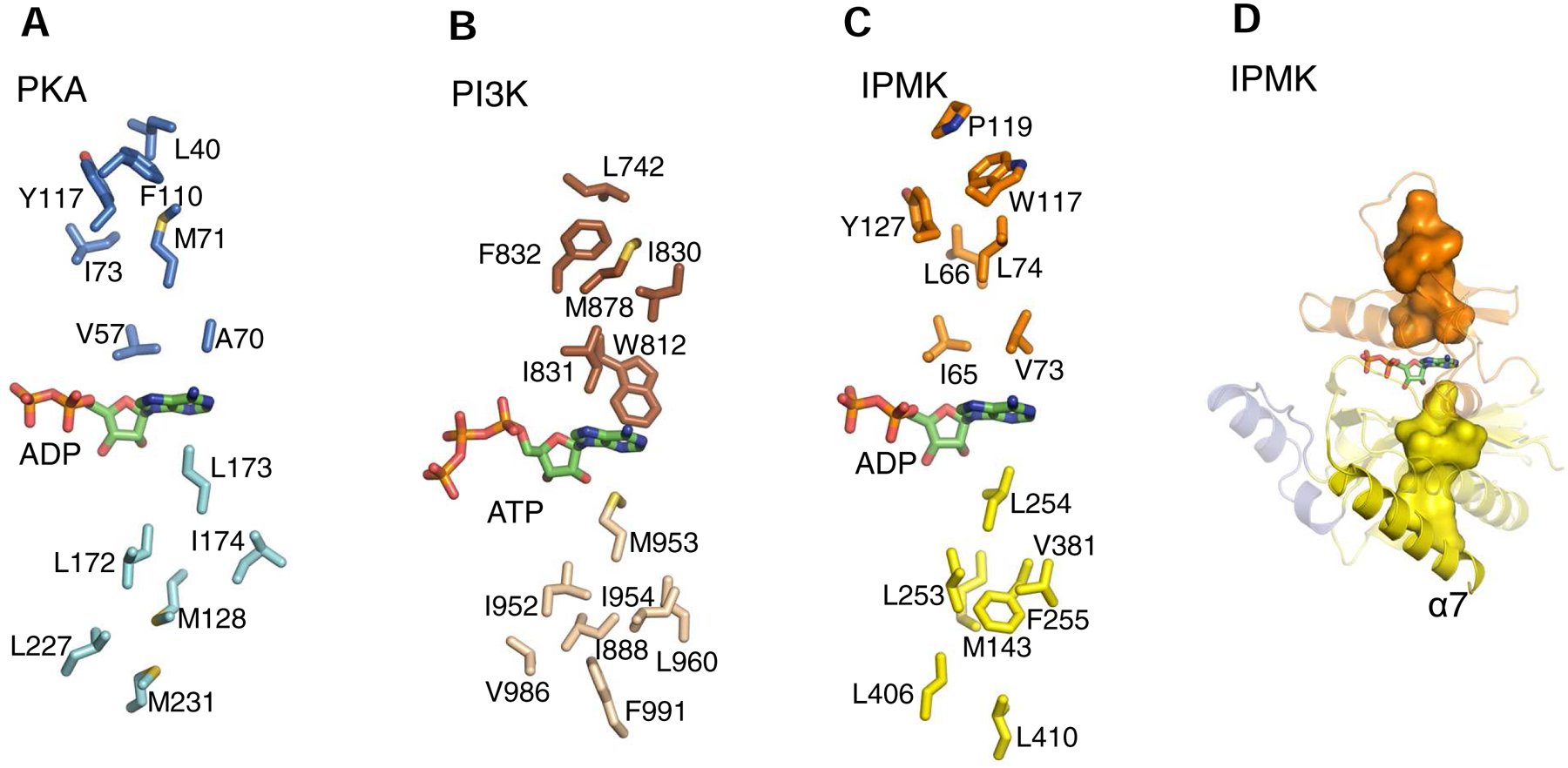
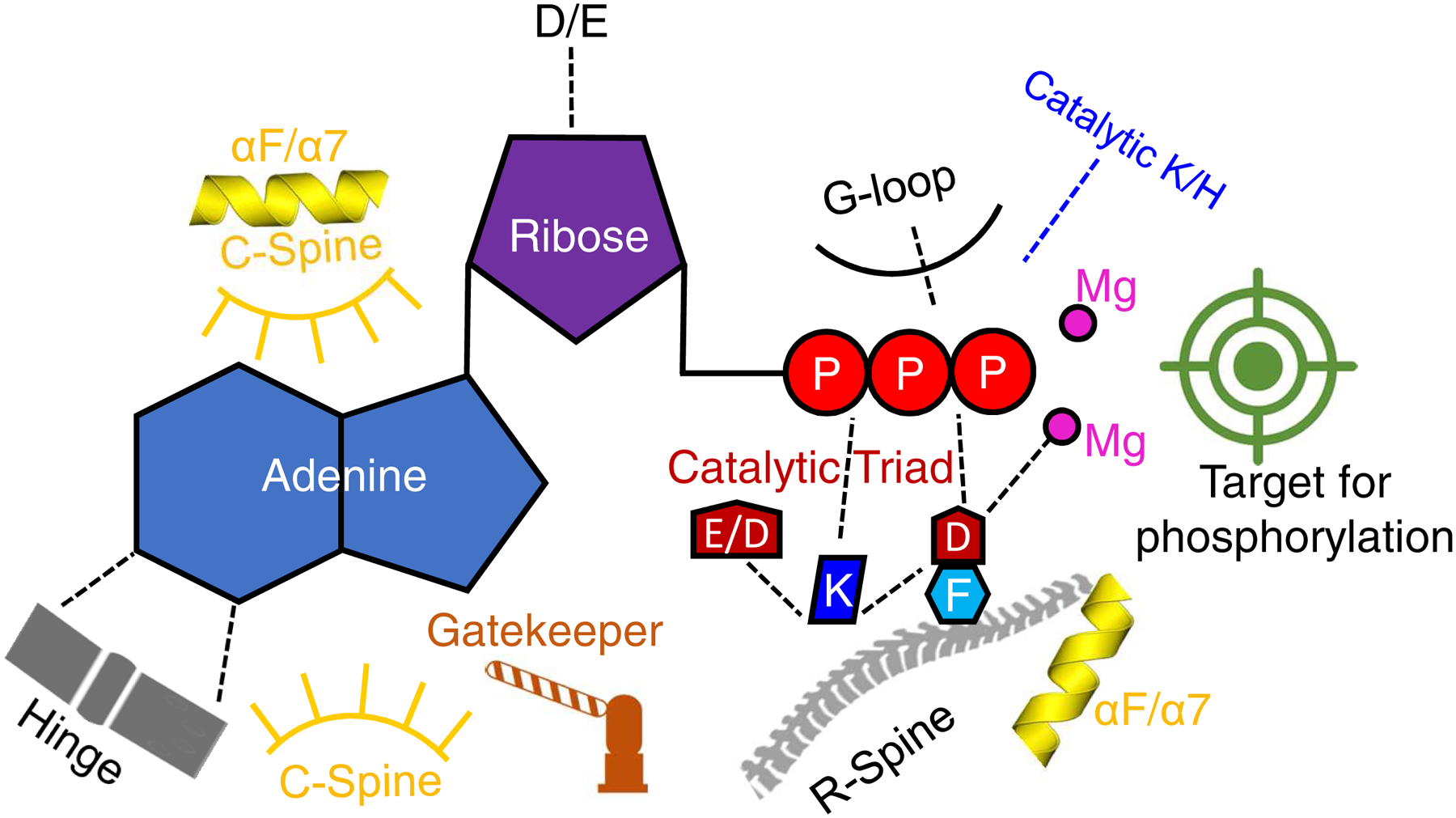
The protein kinase family is characterized by substantial conservation of architectural elements that are required for both ATP binding and phosphotransferase activity. Many of these structural features have also been identified in homologous enzymes that phosphorylate a variety of alternative, non-protein substrates. A comparative structural analysis of these different kinase sub-classes is a portal to a greater understanding of reaction mechanisms, enzyme regulation, inhibitor-development strategies, and superfamily-level evolutionary relationships. To serve such advances, we review structural elements of the protein kinase fold that are conserved in the subfamily of inositol phosphate kinases (InsPKs) that share a PxxxDxKxG catalytic signature: inositol 1,4,5-trisphosphate kinase (IP3K), inositol hexakisphosphate kinase (IP6K), and inositol polyphosphate multikinase (IPMK). We describe conservation of the fundamental two-lobe kinase architecture: an N-lobe constructed upon an anti-parallel β-strand scaffold, which is coupled to a largely helical C-lobe by a single, adenine-binding hinge. This equivalency also includes a G-loop that embraces the β/γ-phosphates of ATP, a transition-state stabilizing residue (Lys/His), and a Mg-positioning aspartate residue within a catalytic triad. Furthermore, we expand this list of conserved structural features to include some not previously identified in InsPKs: a 'gatekeeper' residue in the N-lobe, and an 'αF'-like helix in the C-lobe that anchors two structurally-stabilizing, hydrophobic spines, formed from non-consecutive residues that span the two lobes. We describe how this wide-ranging structural homology can be exploited to develop lead inhibitors of IP6K and IPMK, by using strategies similar to those that have generated ATP-competing inhibitors of protein-kinases. We provide several examples to illustrate how such an approach could benefit human health.
Copyright © 2018. Published by Elsevier Ltd.
Conflict of interest statement
Statement
The authors declare there is no conflict of interest.
Figures
Similar articles
-
Structural features of human inositol phosphate multikinase rationalize its inositol phosphate kinase and phosphoinositide 3-kinase activities.J Biol Chem. 2017 Nov 3;292(44):18192-18202. doi: 10.1074/jbc.M117.801845. Epub 2017 Sep 7. J Biol Chem. 2017. PMID: 28882892 Free PMC article.
-
Inhibition of Inositol Polyphosphate Kinases by Quercetin and Related Flavonoids: A Structure-Activity Analysis.J Med Chem. 2019 Feb 14;62(3):1443-1454. doi: 10.1021/acs.jmedchem.8b01593. Epub 2019 Jan 25. J Med Chem. 2019. PMID: 30624931 Free PMC article.
-
IP6K structure and the molecular determinants of catalytic specificity in an inositol phosphate kinase family.Nat Commun. 2014 Jun 24;5:4178. doi: 10.1038/ncomms5178. Nat Commun. 2014. PMID: 24956979 Free PMC article.
-
The Expanding Significance of Inositol Polyphosphate Multikinase as a Signaling Hub.Mol Cells. 2017 May 31;40(5):315-321. doi: 10.14348/molcells.2017.0066. Epub 2017 May 29. Mol Cells. 2017. PMID: 28554203 Free PMC article. Review.
-
Pharmacological tools to investigate inositol polyphosphate kinases - Enzymes of increasing therapeutic relevance.Adv Biol Regul. 2022 Jan;83:100836. doi: 10.1016/j.jbior.2021.100836. Epub 2021 Nov 9. Adv Biol Regul. 2022. PMID: 34802993 Review.
Cited by
-
Structures of Fission Yeast Inositol Pyrophosphate Kinase Asp1 in Ligand-Free, Substrate-Bound, and Product-Bound States.mBio. 2022 Dec 20;13(6):e0308722. doi: 10.1128/mbio.03087-22. Epub 2022 Dec 5. mBio. 2022. PMID: 36468882 Free PMC article.
-
Structural analyses of inositol phosphate second messengers bound to signaling effector proteins.Adv Biol Regul. 2020 Jan;75:100667. doi: 10.1016/j.jbior.2019.100667. Epub 2019 Oct 11. Adv Biol Regul. 2020. PMID: 31648945 Free PMC article. Review.
-
Synthesis and biological evaluation of flavonoid-based IP6K2 inhibitors.J Enzyme Inhib Med Chem. 2023 Dec;38(1):2193866. doi: 10.1080/14756366.2023.2193866. J Enzyme Inhib Med Chem. 2023. PMID: 37013838 Free PMC article.
-
Development of Novel IP6K Inhibitors for the Treatment of Obesity and Obesity-Induced Metabolic Dysfunctions.J Med Chem. 2022 May 12;65(9):6869-6887. doi: 10.1021/acs.jmedchem.2c00220. Epub 2022 Apr 25. J Med Chem. 2022. PMID: 35467861 Free PMC article.
-
Activities and Structure-Function Analysis of Fission Yeast Inositol Pyrophosphate (IPP) Kinase-Pyrophosphatase Asp1 and Its Impact on Regulation of pho1 Gene Expression.mBio. 2022 Jun 28;13(3):e0103422. doi: 10.1128/mbio.01034-22. Epub 2022 May 10. mBio. 2022. PMID: 35536002 Free PMC article.
References
-
- Abel K, Anderson RA, Shears SB, 2002. Phosphatidylinositol and inositol phosphate metabolism. J. Cell Sci 114, 2207–2208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
