

Cell Membranes Resist Flow
- PMID: 30392960
- PMCID: PMC6541487
- DOI: 10.1016/j.cell.2018.09.054
Cell Membranes Resist Flow
Abstract
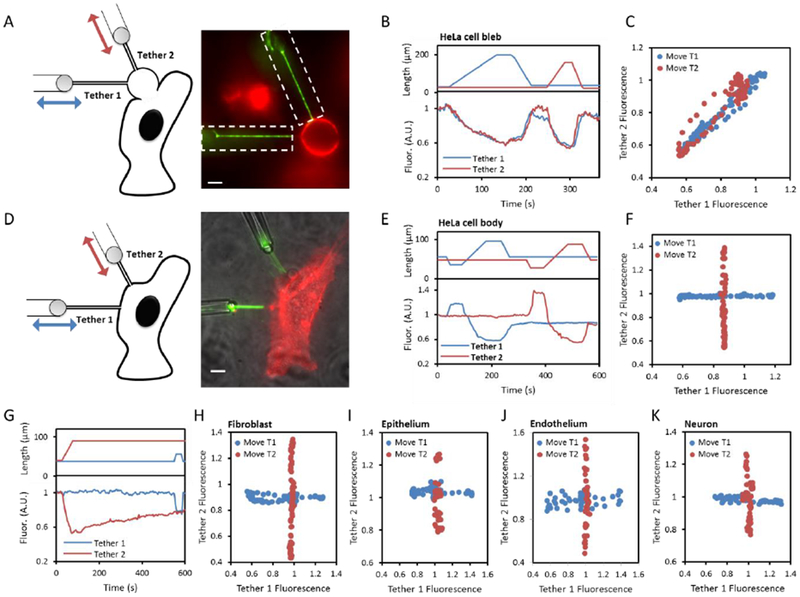
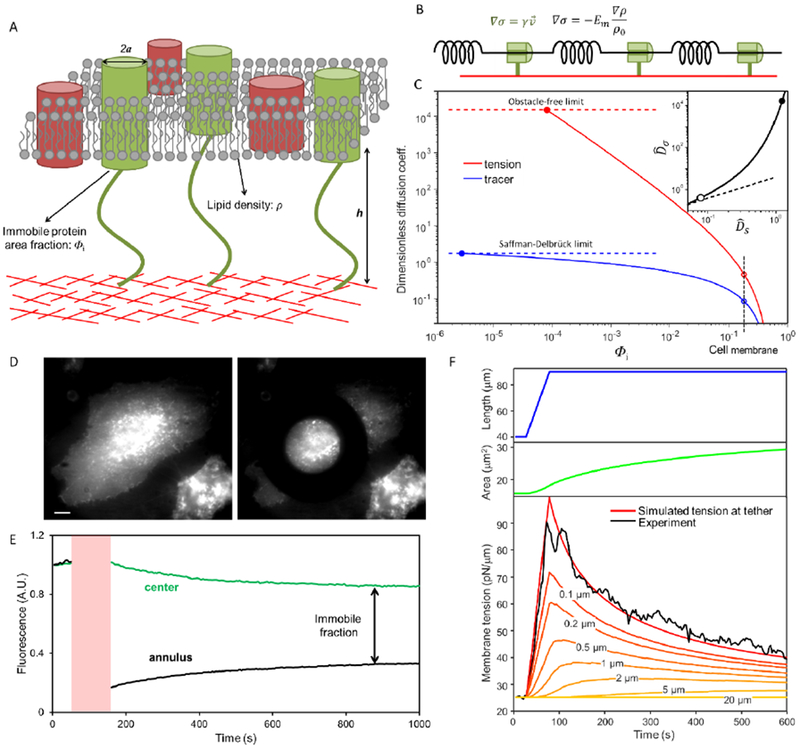
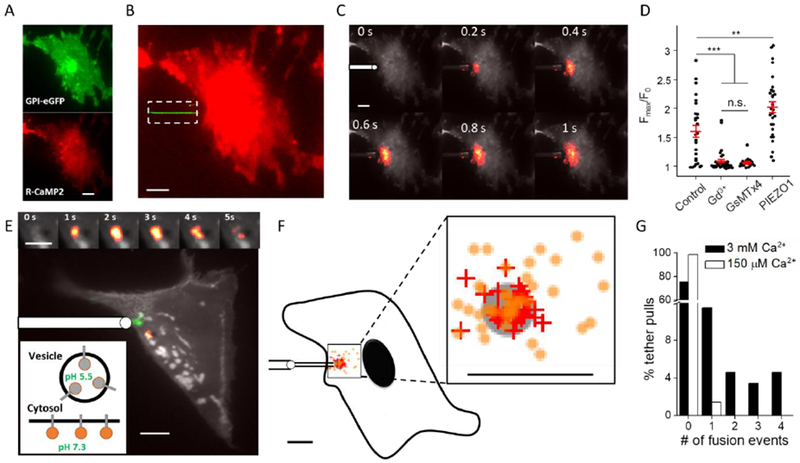
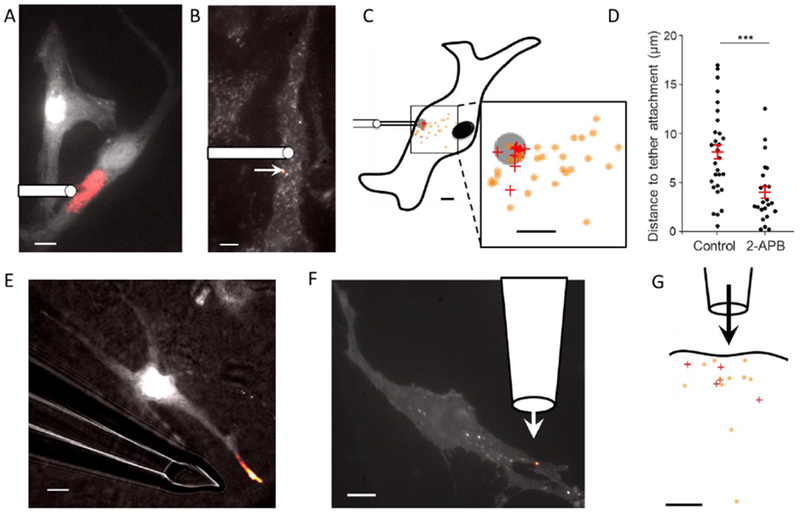
The fluid-mosaic model posits a liquid-like plasma membrane, which can flow in response to tension gradients. It is widely assumed that membrane flow transmits local changes in membrane tension across the cell in milliseconds, mediating long-range signaling. Here, we show that propagation of membrane tension occurs quickly in cell-attached blebs but is largely suppressed in intact cells. The failure of tension to propagate in cells is explained by a fluid dynamical model that incorporates the flow resistance from cytoskeleton-bound transmembrane proteins. Perturbations to tension propagate diffusively, with a diffusion coefficient Dσ ∼0.024 μm2/s in HeLa cells. In primary endothelial cells, local increases in membrane tension lead only to local activation of mechanosensitive ion channels and to local vesicle fusion. Thus, membrane tension is not a mediator of long-range intracellular signaling, but local variations in tension mediate distinct processes in sub-cellular domains.
Keywords: cell mechanics; membrane signaling; membrane tension; porous media; rheology.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARITION OF INTERESTS
The authors declare no competing financial interests.
Figures
Comment in
-
Membrane Mechanics in Living Cells.Dev Cell. 2019 Jan 7;48(1):15-16. doi: 10.1016/j.devcel.2018.12.011. Dev Cell. 2019. PMID: 30620900
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
