

Abnormal Locus Coeruleus Sleep Activity Alters Sleep Signatures of Memory Consolidation and Impairs Place Cell Stability and Spatial Memory
- PMID: 30393040
- PMCID: PMC7556718
- DOI: 10.1016/j.cub.2018.09.054
Abnormal Locus Coeruleus Sleep Activity Alters Sleep Signatures of Memory Consolidation and Impairs Place Cell Stability and Spatial Memory
Abstract
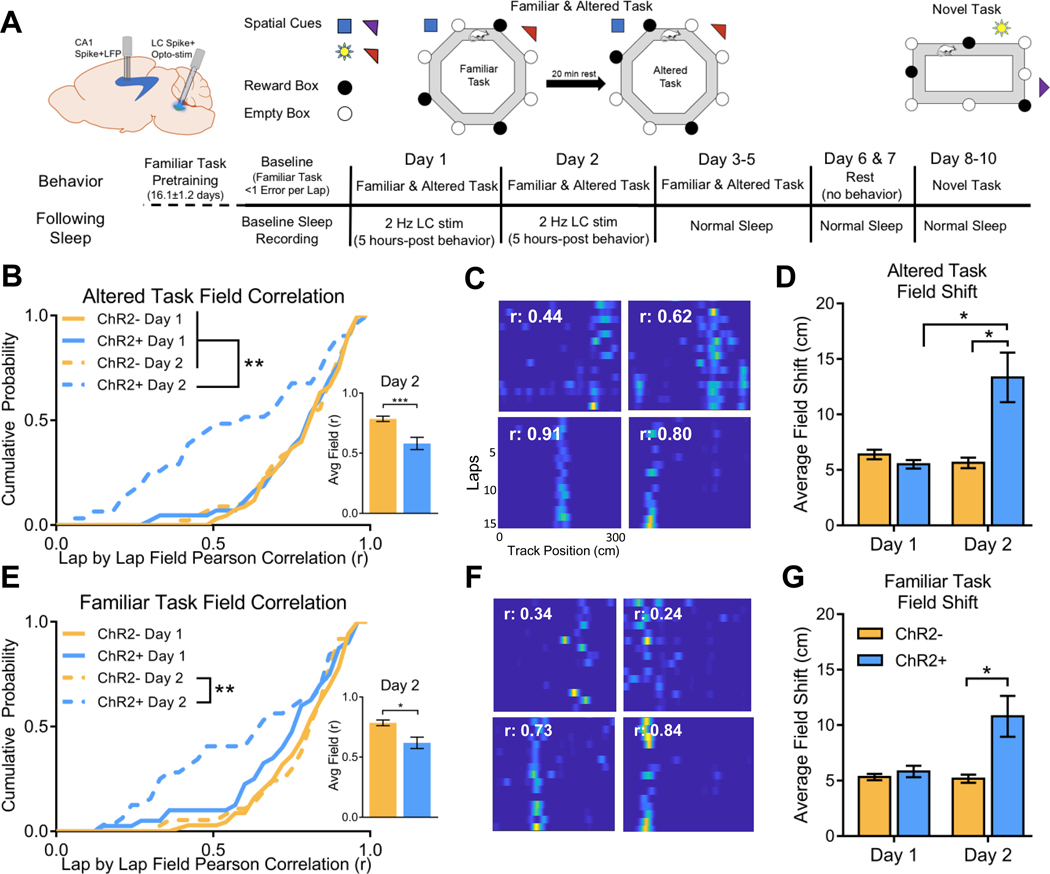
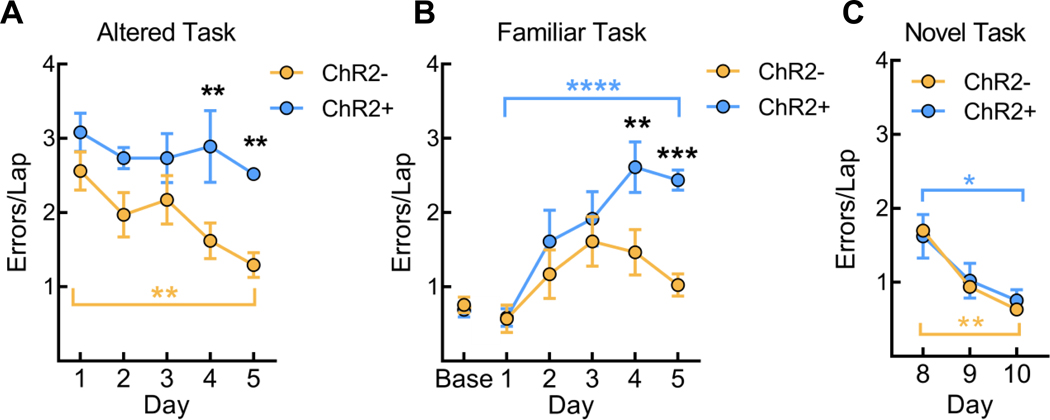
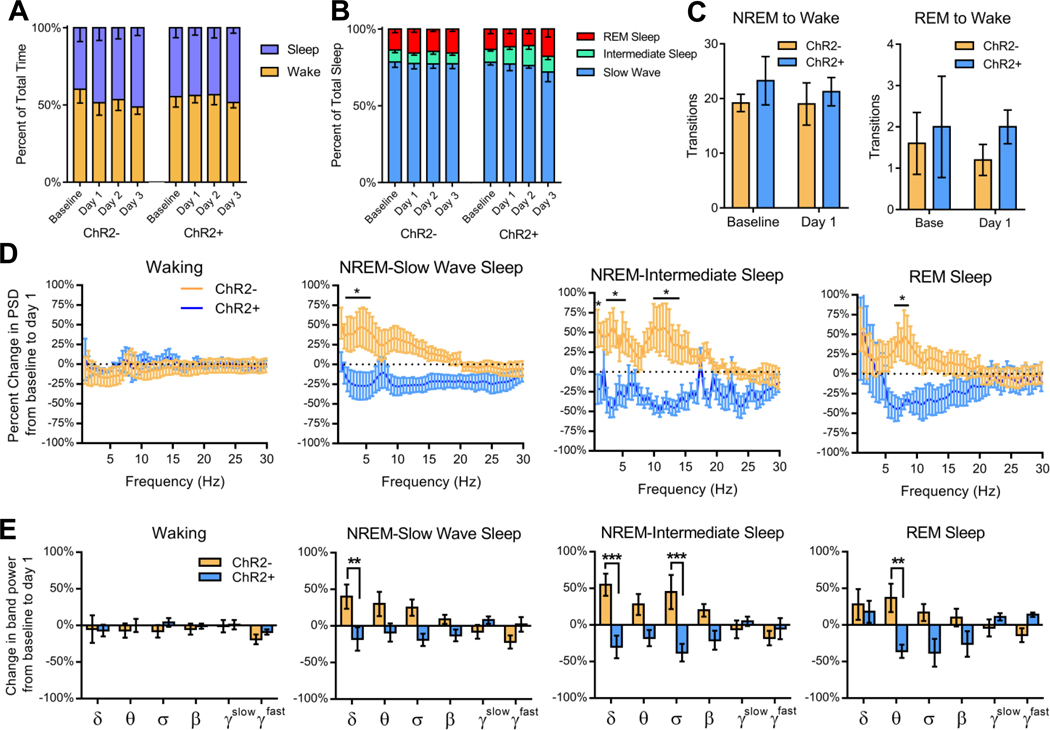
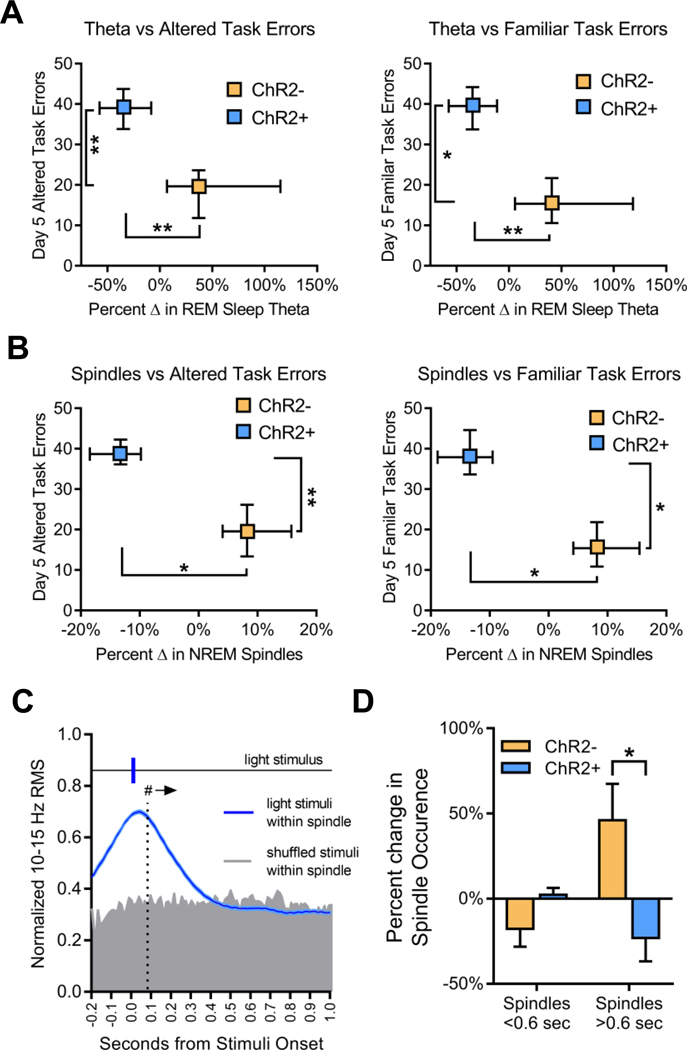
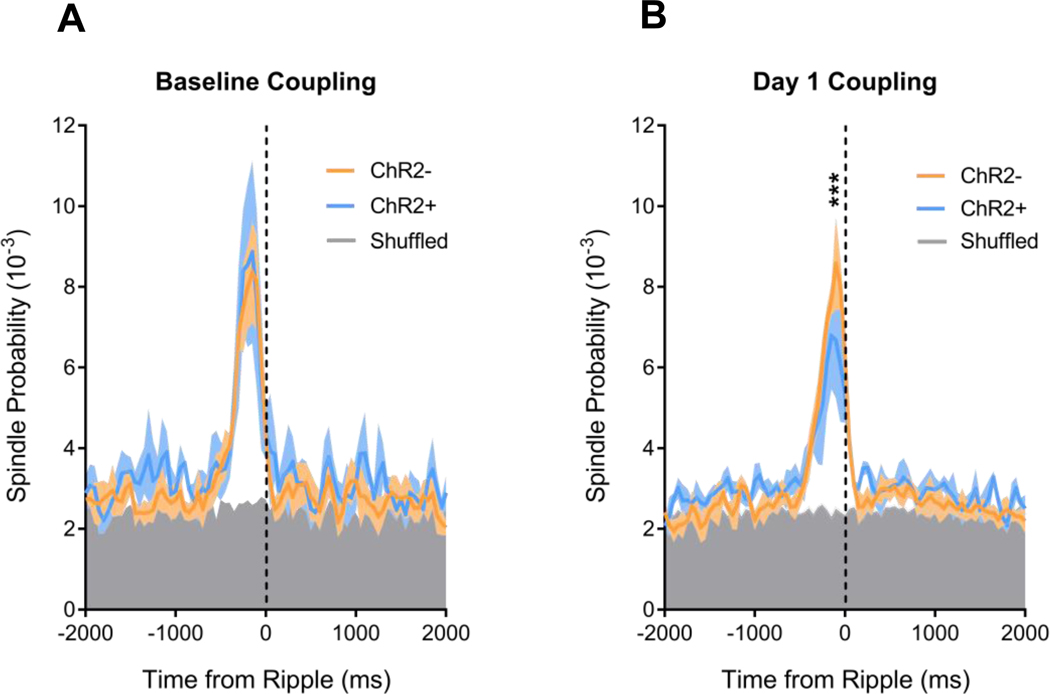
Sleep is critical for proper memory consolidation. The locus coeruleus (LC) releases norepinephrine throughout the brain except when the LC falls silent throughout rapid eye movement (REM) sleep and prior to each non-REM (NREM) sleep spindle. We hypothesize that these transient LC silences allow the synaptic plasticity that is necessary to incorporate new information into pre-existing memory circuits. We found that spontaneous LC activity within sleep spindles triggers a decrease in spindle power. By optogenetically stimulating norepinephrine-containing LC neurons at 2 Hz during sleep, we reduced sleep spindle occurrence, as well as NREM delta power and REM theta power, without causing arousals or changing sleep amounts. Stimulating the LC during sleep following a hippocampus-dependent food location learning task interfered with consolidation of newly learned locations and reconsolidation of previous locations, disrupting next-day place cell activity. The LC stimulation-induced reduction in NREM sleep spindles, delta, and REM theta and reduced ripple-spindle coupling all correlated with decreased hippocampus-dependent performance on the task. Thus, periods of LC silence during sleep following learning are essential for normal spindle generation, delta and theta power, and consolidation of spatial memories.
Keywords: NREM; REM; hippocampus; locus coeruleus; optogenetic; reconsolidation; ripples; sleep; spindle; theta.
Crown Copyright © 2018. Published by Elsevier Ltd. All rights reserved.
Figures


Similar articles
-
Ripple-triggered stimulation of the locus coeruleus during post-learning sleep disrupts ripple/spindle coupling and impairs memory consolidation.Learn Mem. 2016 Apr 15;23(5):238-48. doi: 10.1101/lm.040923.115. Print 2016 May. Learn Mem. 2016. PMID: 27084931 Free PMC article.
-
Hippocampal theta power pressure builds over non-REM sleep and dissipates within REM sleep episodes.Arch Ital Biol. 2018 Sep 1;156(3):112-126. doi: 10.12871/00039829201833. Arch Ital Biol. 2018. PMID: 30324607 Free PMC article.
-
Fragmentation of Rapid Eye Movement and Nonrapid Eye Movement Sleep without Total Sleep Loss Impairs Hippocampus-Dependent Fear Memory Consolidation.Sleep. 2016 Nov 1;39(11):2021-2031. doi: 10.5665/sleep.6236. Sleep. 2016. PMID: 27568801 Free PMC article.
-
Hijacking of hippocampal-cortical oscillatory coupling during sleep in temporal lobe epilepsy.Epilepsy Behav. 2021 Aug;121(Pt B):106608. doi: 10.1016/j.yebeh.2019.106608. Epub 2019 Nov 15. Epilepsy Behav. 2021. PMID: 31740330 Review.
-
Brain neural patterns and the memory function of sleep.Science. 2021 Oct 29;374(6567):560-564. doi: 10.1126/science.abi8370. Epub 2021 Oct 28. Science. 2021. PMID: 34709916 Free PMC article. Review.
Cited by
-
Experience alters hippocampal and cortical network communication via a KIBRA-dependent mechanism.Cell Rep. 2023 Jun 27;42(6):112662. doi: 10.1016/j.celrep.2023.112662. Epub 2023 Jun 20. Cell Rep. 2023. PMID: 37347662 Free PMC article.
-
Neuromodulation of the mind-wandering brain state: the interaction between neuromodulatory tone, sharp wave-ripples and spontaneous thought.Philos Trans R Soc Lond B Biol Sci. 2021 Feb;376(1817):20190699. doi: 10.1098/rstb.2019.0699. Epub 2020 Dec 14. Philos Trans R Soc Lond B Biol Sci. 2021. PMID: 33308063 Free PMC article. Review.
-
Sleep microstructure organizes memory replay.Nature. 2025 Jan;637(8048):1161-1169. doi: 10.1038/s41586-024-08340-w. Epub 2025 Jan 1. Nature. 2025. PMID: 39743590 Free PMC article.
-
WaveSleepNet: An interpretable deep convolutional neural network for the continuous classification of mouse sleep and wake.J Neurosci Methods. 2021 Aug 1;360:109224. doi: 10.1016/j.jneumeth.2021.109224. Epub 2021 May 28. J Neurosci Methods. 2021. PMID: 34052291 Free PMC article.
-
Do your troubles today seem further away than yesterday? On sleep's role in mitigating the blushing response to a reactivated embarrassing episode.Sleep. 2022 Nov 9;45(11):zsac220. doi: 10.1093/sleep/zsac220. Sleep. 2022. PMID: 36130113 Free PMC article.
References
-
- Siapas AG, and Wilson MA (1998). Coordinated Interactions between Hippocampal Ripples and Cortical Spindles during Slow-Wave Sleep. Neuron 21, 1123–1128. - PubMed
-
- Ji D, and Wilson MA (2007). Coordinated memory replay in the visual cortex and hippocampus during sleep. Nat. Neurosci. 10, 100–107. - PubMed
-
- Sara SJ (2009). The locus coeruleus and noradrenergic modulation of cognition. Nat. Rev. Neurosci. 10, 211–223. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
