

Synthetic Fluorogenic Peptides Reveal Dynamic Substrate Specificity of Depalmitoylases
- PMID: 30393067
- PMCID: PMC6338520
- DOI: 10.1016/j.chembiol.2018.10.005
Synthetic Fluorogenic Peptides Reveal Dynamic Substrate Specificity of Depalmitoylases
Abstract
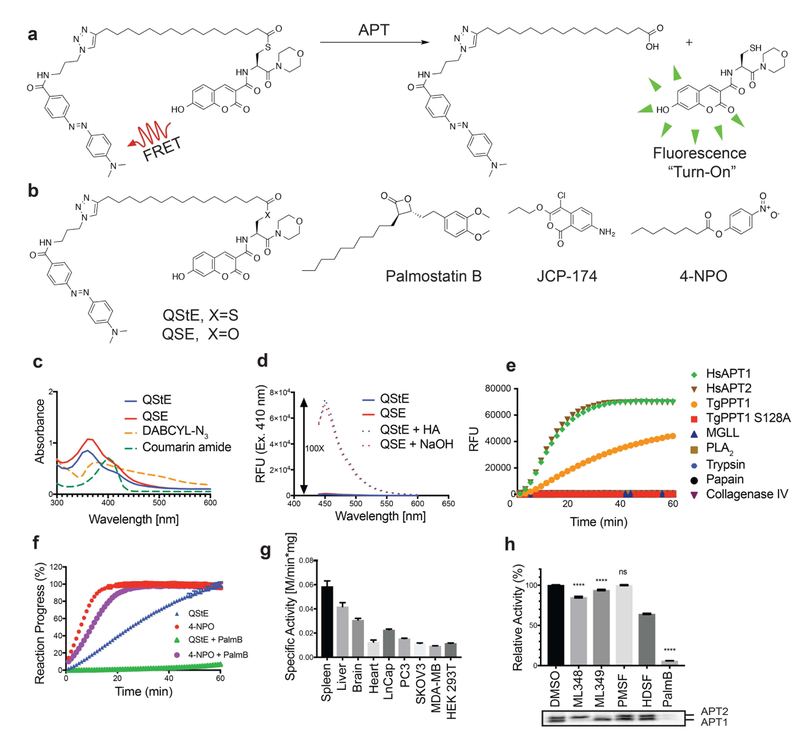
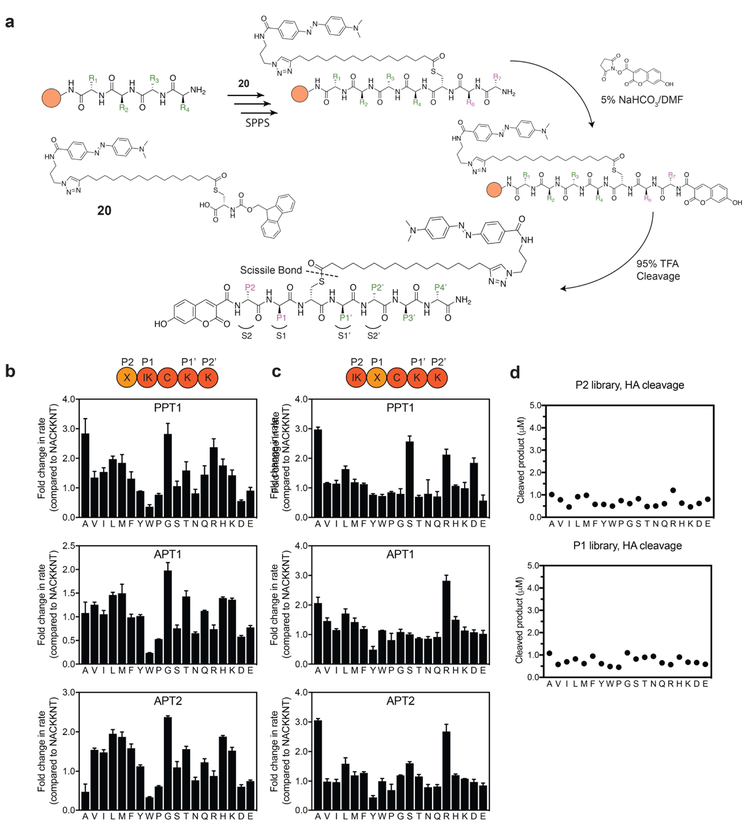
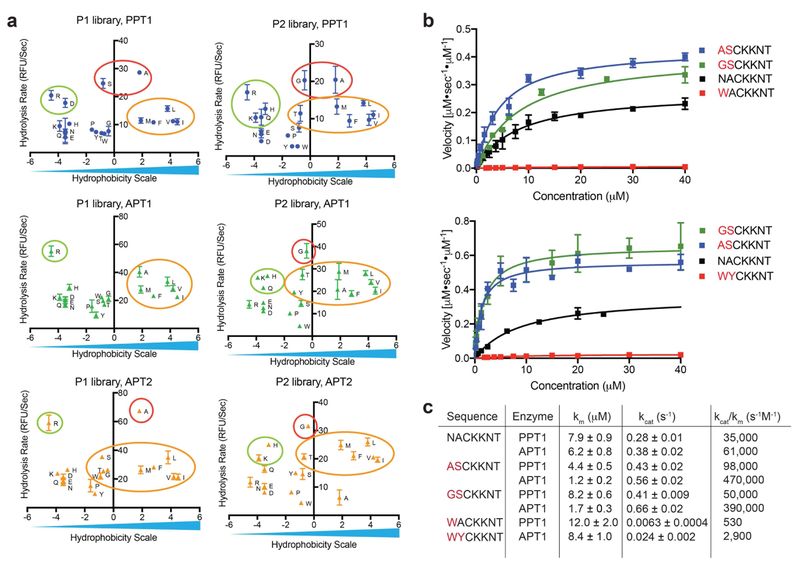
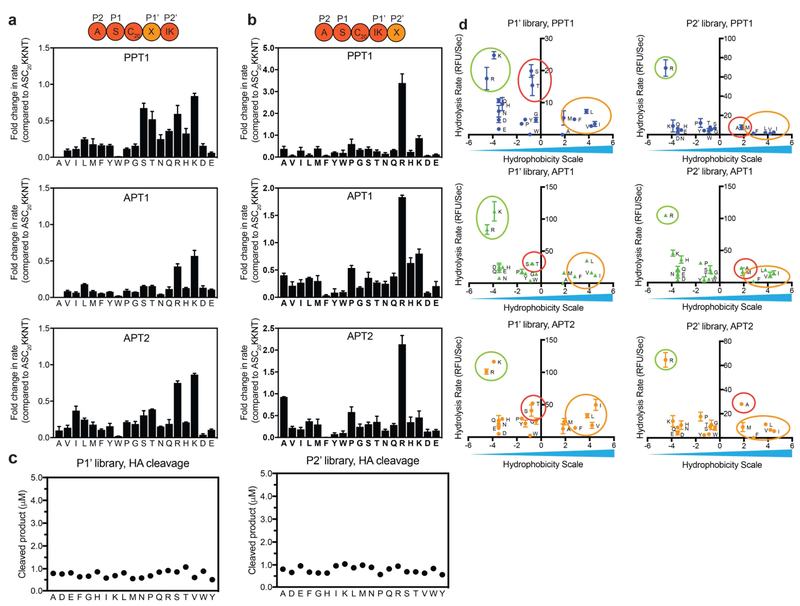
Palmitoylation is a post-translational modification involving the thioesterification of cysteine residues with a 16-carbon-saturated fatty acid. Little is known about rates of depalmitoylation or the parameters that dictate these rates. Here we report a modular strategy to synthesize quenched fluorogenic substrates for the specific detection of depalmitoylase activity and for mapping the substrate specificity of individual depalmitoylases. We demonstrate that human depalmitoylases APT1 and APT2, and TgPPT1 from the parasite Toxoplasma gondii, have distinct specificities that depend on amino acid residues distal to the palmitoyl cysteine. This information informs the design of optimal and non-optimal substrates as well as isoform-selective substrates to detect the activity of a specific depalmitoylase in complex proteomes. In addition to providing tools for studying depalmitoylases, our findings identify a previously unrecognized mechanism for regulating steady-state levels of distinct palmitoylation sites by sequence-dependent control of depalmitoylation rates.
Keywords: chemical probes; combinatorial libraries; depalmitoylases; dynamic palmitoylation; fluorogenic peptides; substrate specificity; thioesterases.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Figures
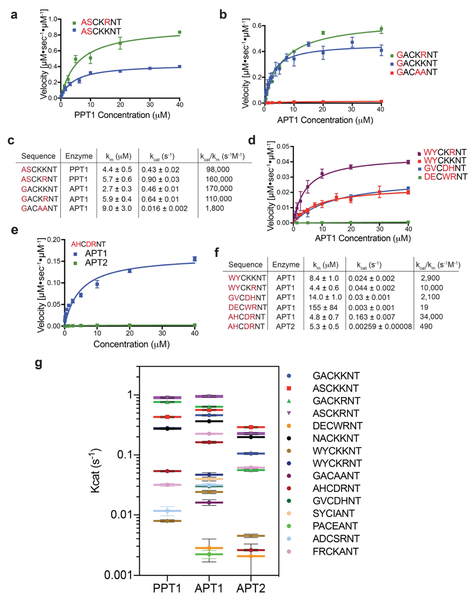
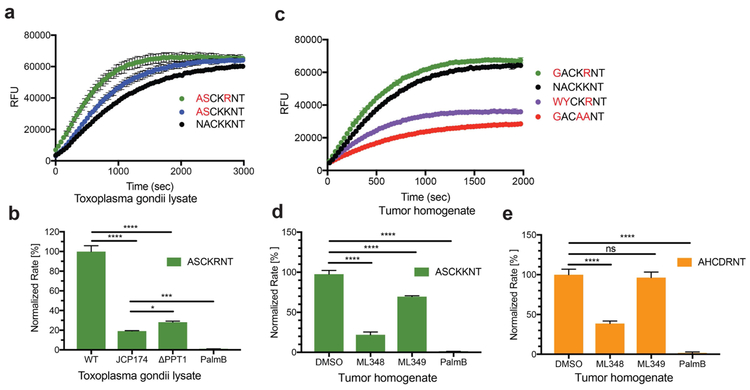
Comment in
-
Probing Substrate Preferences of Depalmitoylases.Cell Chem Biol. 2019 Jan 17;26(1):3-6. doi: 10.1016/j.chembiol.2018.12.008. Cell Chem Biol. 2019. PMID: 30658110
Similar articles
-
Activity-Based Sensing of S-Depalmitoylases: Chemical Technologies and Biological Discovery.Acc Chem Res. 2019 Nov 19;52(11):3029-3038. doi: 10.1021/acs.accounts.9b00354. Epub 2019 Oct 2. Acc Chem Res. 2019. PMID: 31577124 Free PMC article. Review.
-
Active and dynamic mitochondrial S-depalmitoylation revealed by targeted fluorescent probes.Nat Commun. 2018 Jan 23;9(1):334. doi: 10.1038/s41467-017-02655-1. Nat Commun. 2018. PMID: 29362370 Free PMC article.
-
Protein depalmitoylases.Crit Rev Biochem Mol Biol. 2018 Feb;53(1):83-98. doi: 10.1080/10409238.2017.1409191. Epub 2017 Dec 14. Crit Rev Biochem Mol Biol. 2018. PMID: 29239216 Free PMC article. Review.
-
A Fluorescent Probe with Improved Water Solubility Permits the Analysis of Protein S-Depalmitoylation Activity in Live Cells.Biochemistry. 2018 Jan 16;57(2):221-225. doi: 10.1021/acs.biochem.7b00835. Epub 2017 Oct 18. Biochemistry. 2018. PMID: 29023093 Free PMC article.
-
Dynamic palmitoylation links cytosol-membrane shuttling of acyl-protein thioesterase-1 and acyl-protein thioesterase-2 with that of proto-oncogene H-ras product and growth-associated protein-43.J Biol Chem. 2013 Mar 29;288(13):9112-25. doi: 10.1074/jbc.M112.421073. Epub 2013 Feb 8. J Biol Chem. 2013. PMID: 23396970 Free PMC article.
Cited by
-
Depalmitoylation and cell physiology: APT1 as a mediator of metabolic signals.Am J Physiol Cell Physiol. 2024 Apr 1;326(4):C1034-C1041. doi: 10.1152/ajpcell.00542.2023. Epub 2024 Feb 12. Am J Physiol Cell Physiol. 2024. PMID: 38344800 Free PMC article. Review.
-
Glycosylation Analysis of Feline Small Intestine Following Toxoplasma gondii Infection.Animals (Basel). 2022 Oct 20;12(20):2858. doi: 10.3390/ani12202858. Animals (Basel). 2022. PMID: 36290246 Free PMC article.
-
Activity-Based Sensing of S-Depalmitoylases: Chemical Technologies and Biological Discovery.Acc Chem Res. 2019 Nov 19;52(11):3029-3038. doi: 10.1021/acs.accounts.9b00354. Epub 2019 Oct 2. Acc Chem Res. 2019. PMID: 31577124 Free PMC article. Review.
-
Recent advances in activity-based probes (ABPs) and affinity-based probes (AfBPs) for profiling of enzymes.Chem Sci. 2021 May 18;12(24):8288-8310. doi: 10.1039/d1sc01359a. Chem Sci. 2021. PMID: 34221311 Free PMC article. Review.
-
Mechanisms and functions of protein S-acylation.Nat Rev Mol Cell Biol. 2024 Jun;25(6):488-509. doi: 10.1038/s41580-024-00700-8. Epub 2024 Feb 14. Nat Rev Mol Cell Biol. 2024. PMID: 38355760 Free PMC article. Review.
References
-
- Adibekian A, Martin BR, Chang JW, Hsu KL, Tsuboi K, Bachovchin DA, Speers AE, Brown SJ, Spicer T, Fernandez-Vega V, et al. (2010). Characterization of a Selective, Reversible Inhibitor of Lysophospholipase 2 (LYPLA2) In Probe Reports from the NIH Molecular Libraries Program (Bethesda (MD)). - PubMed
-
- Baker TL, Zheng H, Walker J, Coloff JL, and Buss JE (2003). Distinct rates of palmitate turnover on membrane-bound cellular and oncogenic H-ras. J Biol Chem 278, 19292–19300. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
