

The role of 3' end uridylation in RNA metabolism and cellular physiology
- PMID: 30397107
- PMCID: PMC6232591
- DOI: 10.1098/rstb.2018.0171
The role of 3' end uridylation in RNA metabolism and cellular physiology
Abstract
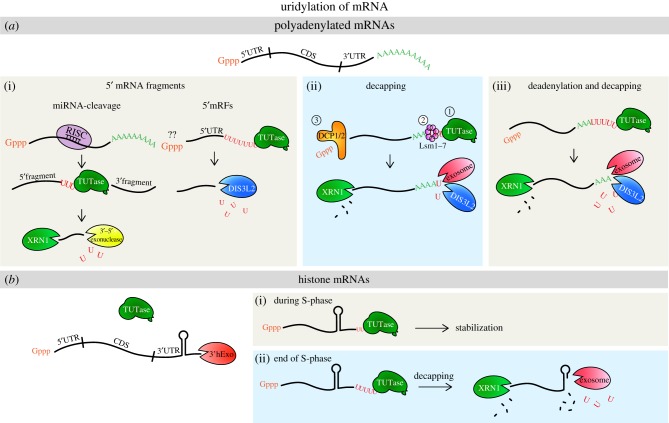
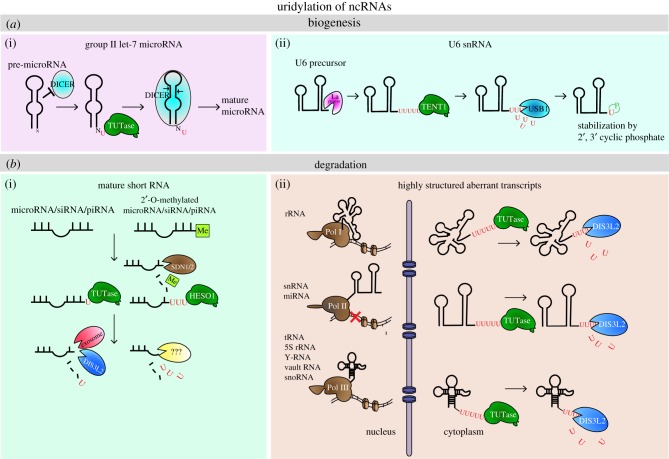
Most eukaryotic RNAs are posttranscriptionally modified. The majority of modifications promote RNA maturation, others may regulate function and stability. The 3' terminal non-templated oligouridylation is a widespread modification affecting many cellular RNAs at some stage of their life cycle. It has diverse roles in RNA metabolism. The most prevalent is the regulation of stability and quality control. On the cellular and organismal level, it plays a critical role in a number of pathways, such as cell cycle regulation, cell death, development or viral infection. Defects in uridylation have been linked to several diseases. This review summarizes the current knowledge about the role of the 3' terminal oligo(U)-tailing in biology of various RNAs in eukaryotes and describes key factors involved in these pathways.This article is part of the theme issue '5' and 3' modifications controlling RNA degradation'.
Keywords: RNA degradation; RNA modification; RNA processing; RNA surveillance; RNA uridylation; tutase.
© 2018 The Author(s).
Conflict of interest statement
We have no competing interests.
Figures
Similar articles
-
Surprises in the 3'-end: 'U' can decide too!FEBS J. 2015 Sep;282(18):3489-99. doi: 10.1111/febs.13377. Epub 2015 Aug 4. FEBS J. 2015. PMID: 26183531 Review.
-
RNA uridylation: a key posttranscriptional modification shaping the coding and noncoding transcriptome.Wiley Interdiscip Rev RNA. 2018 Jan;9(1). doi: 10.1002/wrna.1440. Epub 2017 Oct 5. Wiley Interdiscip Rev RNA. 2018. PMID: 28984054 Review.
-
Polyuridylation in Eukaryotes: A 3'-End Modification Regulating RNA Life.Biomed Res Int. 2015;2015:968127. doi: 10.1155/2015/968127. Epub 2015 May 11. Biomed Res Int. 2015. PMID: 26078976 Free PMC article. Review.
-
Uridylation: a vital way for cellular RNA surveillance.Yi Chuan. 2022 Jun 20;44(6):449-465. doi: 10.16288/j.yczz.22-094. Yi Chuan. 2022. PMID: 35729094 Review.
-
Widespread RNA 3'-end oligouridylation in mammals.RNA. 2012 Mar;18(3):394-401. doi: 10.1261/rna.029306.111. Epub 2012 Jan 30. RNA. 2012. PMID: 22291204 Free PMC article.
Cited by
-
Uridylation and the SKI complex orchestrate the Calvin cycle of photosynthesis through RNA surveillance of TKL1 in Arabidopsis.Proc Natl Acad Sci U S A. 2022 Sep 20;119(38):e2205842119. doi: 10.1073/pnas.2205842119. Epub 2022 Sep 12. Proc Natl Acad Sci U S A. 2022. PMID: 36095196 Free PMC article.
-
lncRNA BC200 is processed into a stable Alu monomer.RNA. 2024 Oct 16;30(11):1477-1494. doi: 10.1261/rna.080152.124. RNA. 2024. PMID: 39179355
-
Functions and mechanisms of RNA tailing by nucleotidyl transferase proteins in plants.Front Plant Sci. 2024 Oct 15;15:1452347. doi: 10.3389/fpls.2024.1452347. eCollection 2024. Front Plant Sci. 2024. PMID: 39474218 Free PMC article. Review.
-
The regulation of antiviral innate immunity through non-m6A RNA modifications.Front Immunol. 2023 Oct 17;14:1286820. doi: 10.3389/fimmu.2023.1286820. eCollection 2023. Front Immunol. 2023. PMID: 37915585 Free PMC article. Review.
-
Pervasive mRNA uridylation in fission yeast is catalysed by both Cid1 and Cid16 terminal uridyltransferases.PLoS One. 2023 May 23;18(5):e0285576. doi: 10.1371/journal.pone.0285576. eCollection 2023. PLoS One. 2023. PMID: 37220133 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
