

Minimal PAM specificity of a highly similar SpCas9 ortholog
- PMID: 30397647
- PMCID: PMC6200363
- DOI: 10.1126/sciadv.aau0766
Minimal PAM specificity of a highly similar SpCas9 ortholog
Abstract
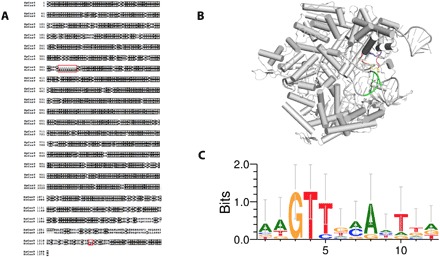
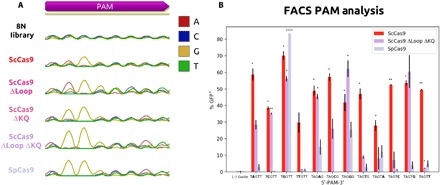
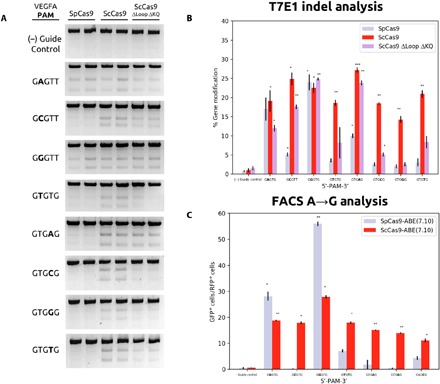
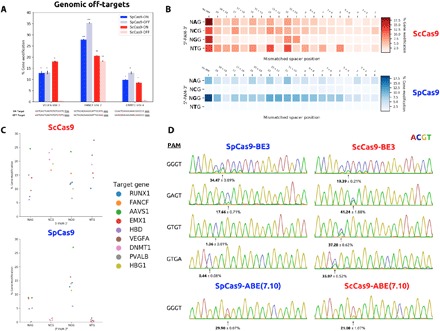
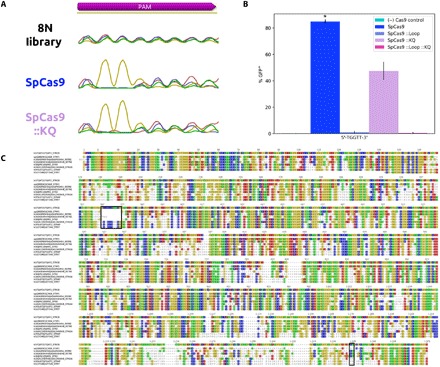
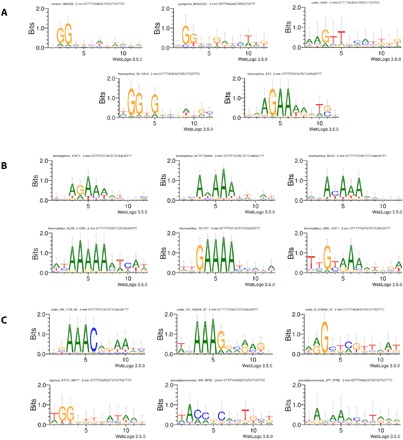
RNA-guided DNA endonucleases of the CRISPR-Cas system are widely used for genome engineering and thus have numerous applications in a wide variety of fields. CRISPR endonucleases, however, require a specific protospacer adjacent motif (PAM) flanking the target site, thus constraining their targetable sequence space. In this study, we demonstrate the natural PAM plasticity of a highly similar, yet previously uncharacterized, Cas9 from Streptococcus canis (ScCas9) through rational manipulation of distinguishing motif insertions. To this end, we report affinity to minimal 5'-NNG-3' PAM sequences and demonstrate the accurate editing capabilities of the ortholog in both bacterial and human cells. Last, we build an automated bioinformatics pipeline, the Search for PAMs by ALignment Of Targets (SPAMALOT), which further explores the microbial PAM diversity of otherwise overlooked Streptococcus Cas9 orthologs. Our results establish that ScCas9 can be used both as an alternative genome editing tool and as a functional platform to discover novel Streptococcus PAM specificities.
Figures
References
-
- Barrangou R., Horvath P., A decade of discovery: CRISPR functions and applications. Nat. Microbiol. 2, 17092 (2017). - PubMed
-
- Mojica F. J., Díez-Villaseñor C., García-Martínez J., Almendros C., Short motif sequences determine the targets of the prokaryotic CRISPR defense system. Microbiology 155, 733–740 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
