

Longitudinal analysis of developmental changes in electroencephalography patterns and sleep-wake states of the neonatal mouse
- PMID: 30399187
- PMCID: PMC6219806
- DOI: 10.1371/journal.pone.0207031
Longitudinal analysis of developmental changes in electroencephalography patterns and sleep-wake states of the neonatal mouse
Abstract
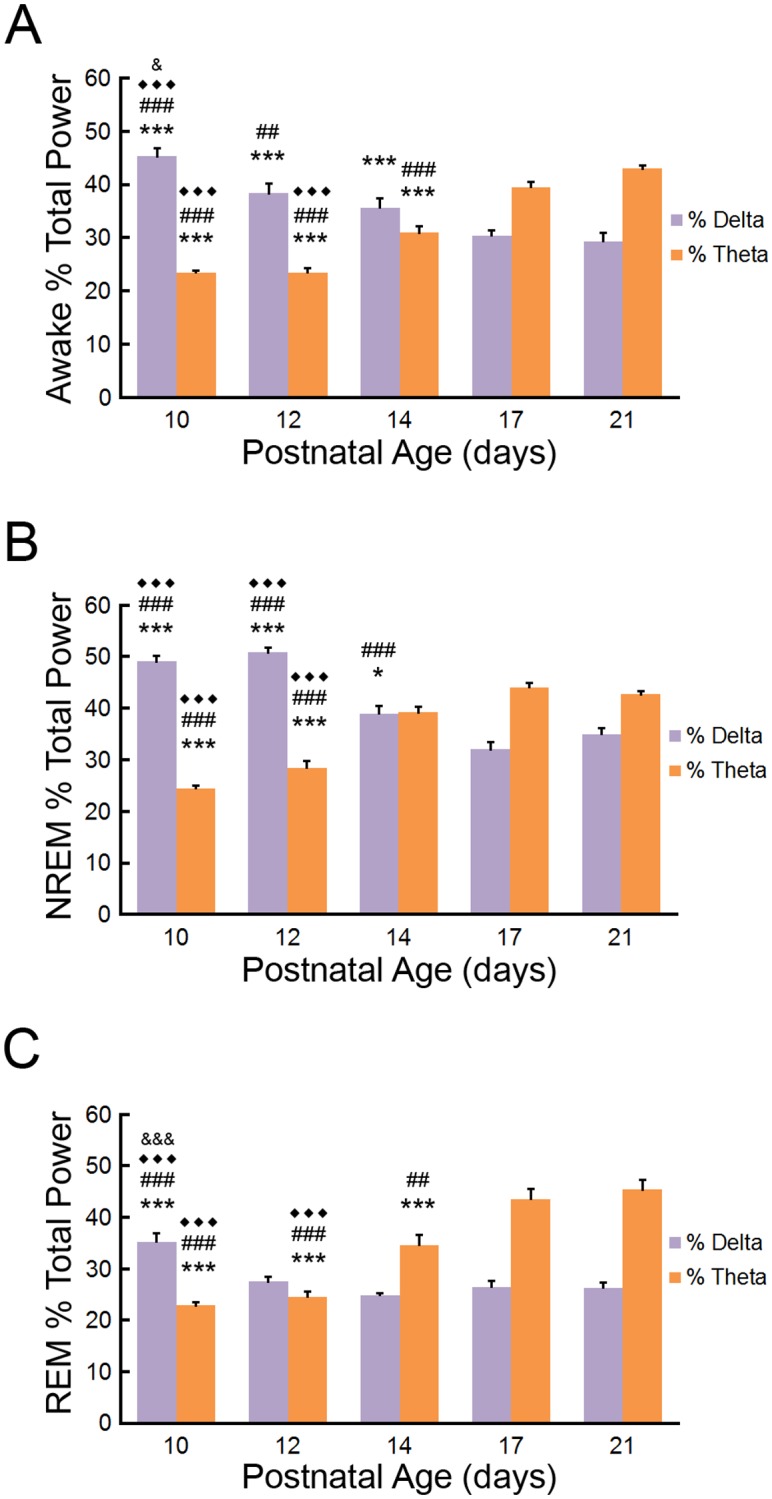
The neonatal brain undergoes rapid maturational changes that facilitate the normal development of the nervous system and also affect the pathological response to brain injury. Electroencephalography (EEG) and analysis of sleep-wake vigilance states provide important insights into the function of the normal and diseased immature brain. While developmental changes in EEG and vigilance states are well-described in people, less is known about the normal maturational properties of rodent EEG, including the emergence and evolution of sleep-awake vigilance states. In particular, a number of developmental EEG studies have been performed in rats, but there is limited comparable research in neonatal mice, especially as it pertains to longitudinal EEG studies performed within the same mouse. In this study, we have attempted to provide a relatively comprehensive assessment of developmental changes in EEG background activity and vigilance states in wild-type mice from postnatal days 9-21. A novel EEG and EMG method allowed serial recording from the same mouse pups. EEG continuity and power and vigilance states were analyzed by quantitative assessment and fast Fourier transforms. During this developmental period, we demonstrate the timing of maturational changes in EEG background continuity, frequencies, and power and the emergence of identifiable wake, NREM, and REM sleep states. These results should serve as important control data for physiological studies of mouse models of normal brain development and neurological disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
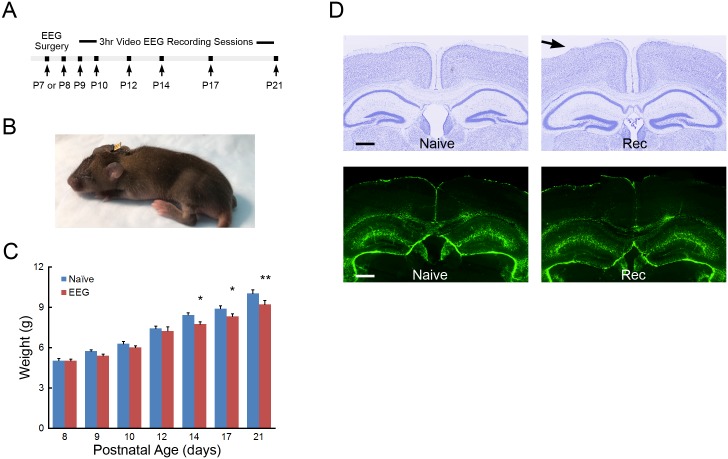
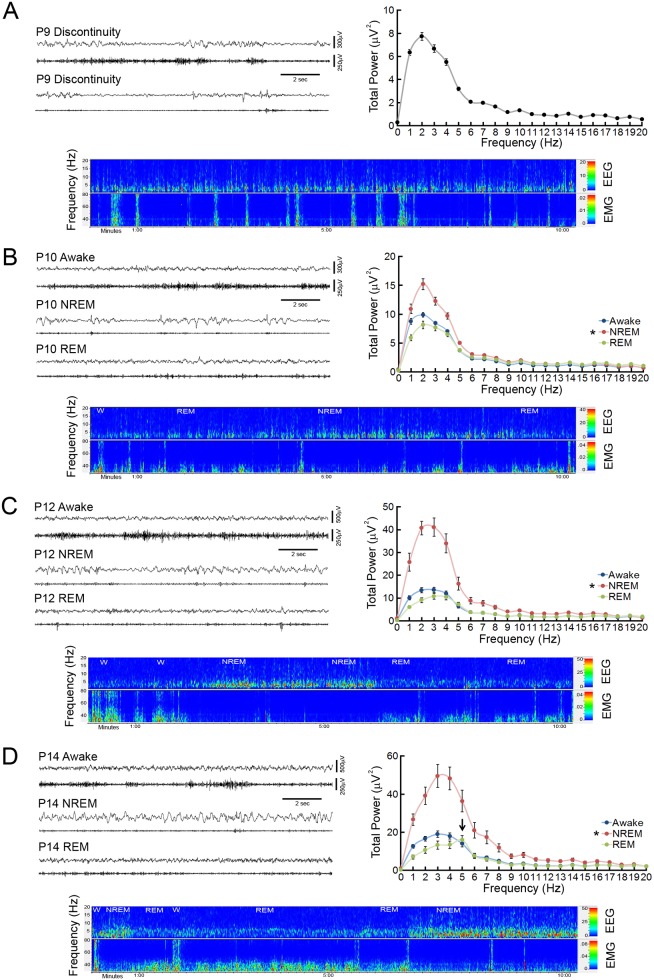
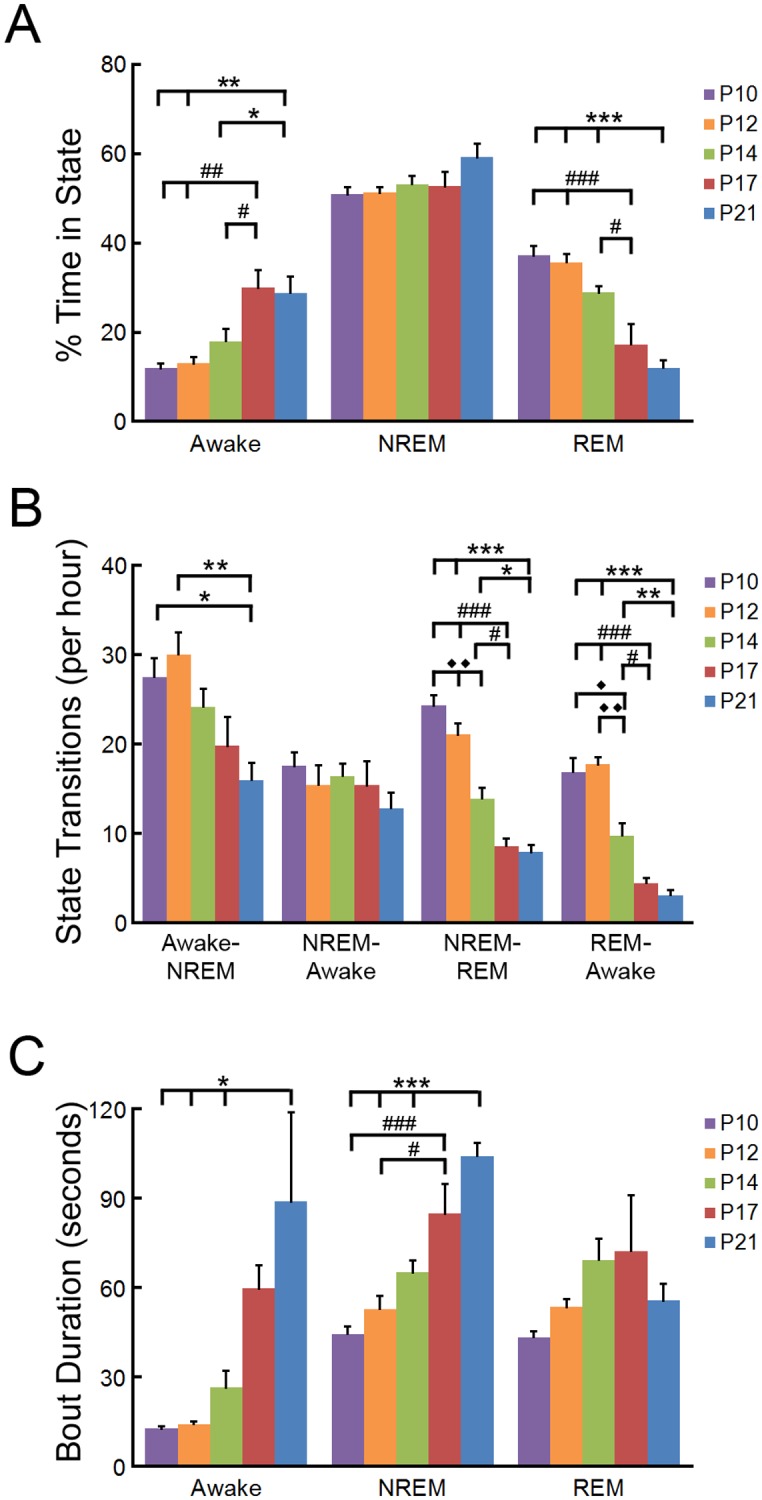
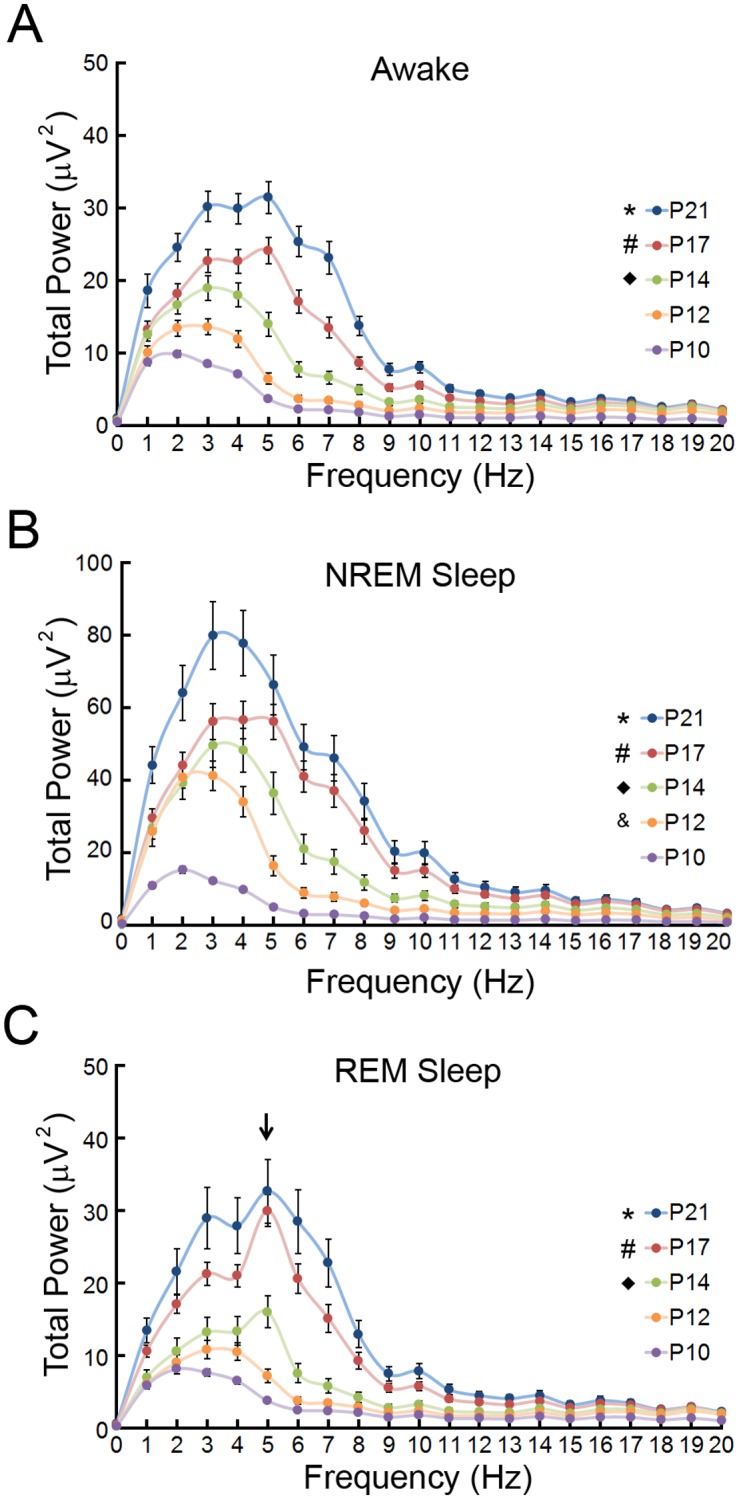
References
-
- Britton JW, Frey LC, Hopp JL, Korb P, Koubeissi MZ, Lievens WE, et al. In: St. Louis EK, Frey LC, editors. Electroencephalography (EEG): An Introductory Text and Atlas of Normal and Abnormal Findings in Adults, Children, and Infants. Chicago: American Epilepsy Society Copyright (c)2016 by American Epilepsy Society; 2016. - PubMed
-
- Andre M, Lamblin MD, d’Allest AM, Curzi-Dascalova L, Moussalli-Salefranque F, SNT T, et al. Electroencephalography in premature and full-term infants. Developmental features and glossary. Neurophysiologie clinique = Clinical neurophysiology. 2010;40(2):59–124. Epub 2010/06/01. 10.1016/j.neucli.2010.02.002 . - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
