

Temperature and photoperiod changes affect cucumber sex expression by different epigenetic regulations
- PMID: 30400867
- PMCID: PMC6220452
- DOI: 10.1186/s12870-018-1490-3
Temperature and photoperiod changes affect cucumber sex expression by different epigenetic regulations
Abstract
Background: Cucumbers (Cucumis sativus) are known for their plasticity in sex expression. DNA methylation status determines gene activity but is susceptible to environmental condition changes. Thus, DNA methylation-based epigenetic regulation may at least partially account for the instability of cucumber sex expression. Do temperature and photoperiod that are the two most important environmental factors have equal effect on cucumber sex expression by similar epigenetic regulation mechanism? To answer this question, we did a two-factor experiment of temperature and photoperiod and generated methylome and transcriptome data from cucumber shoot apices.
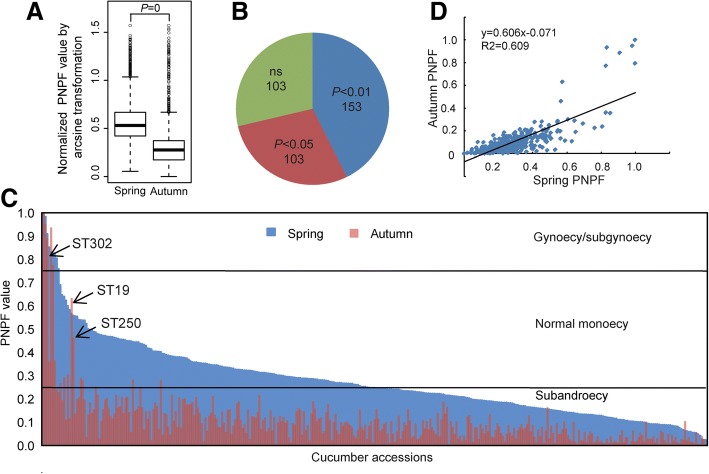
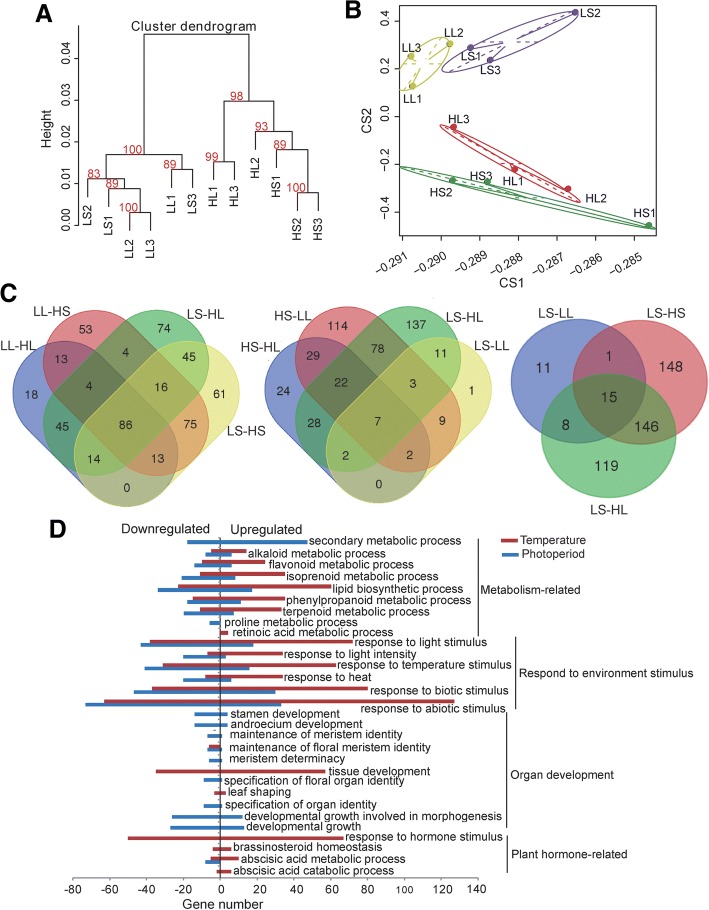
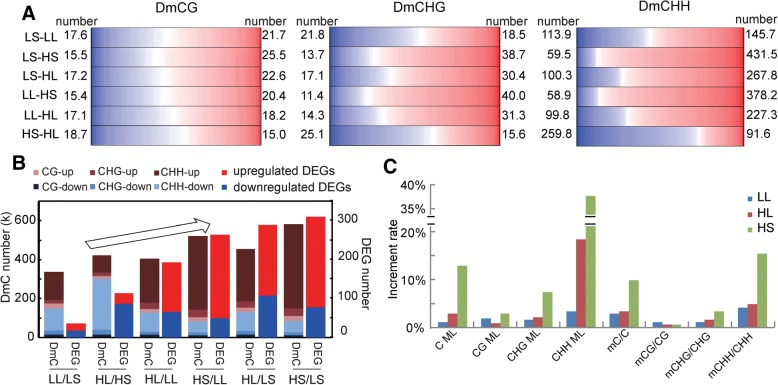
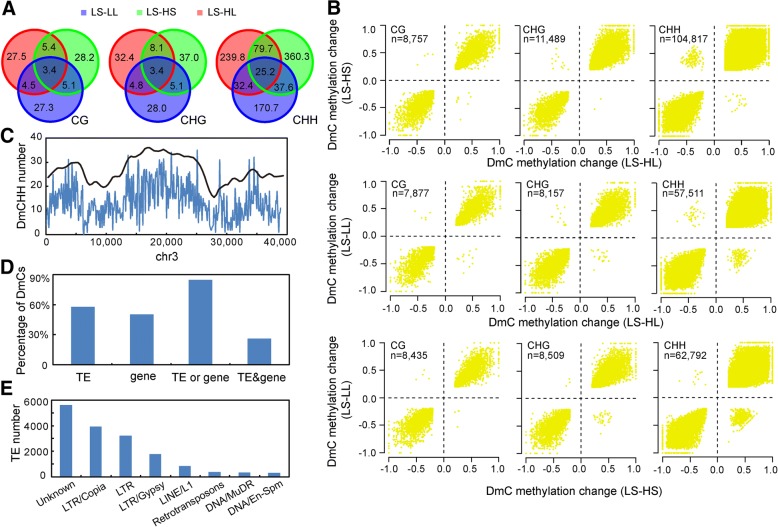
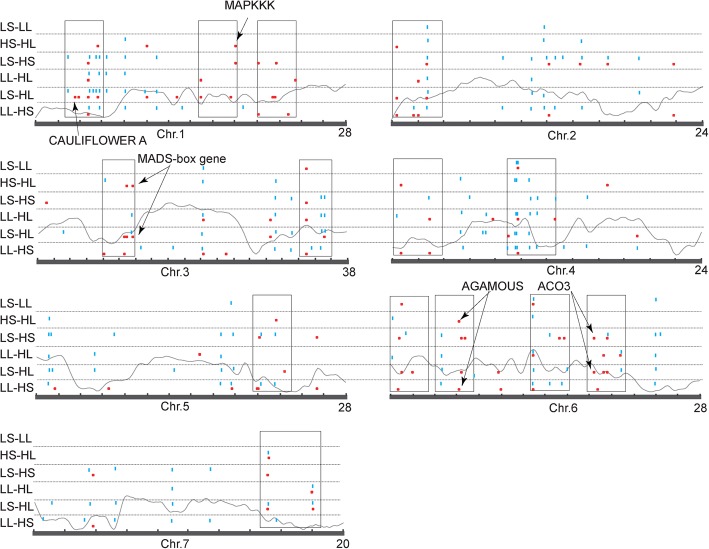
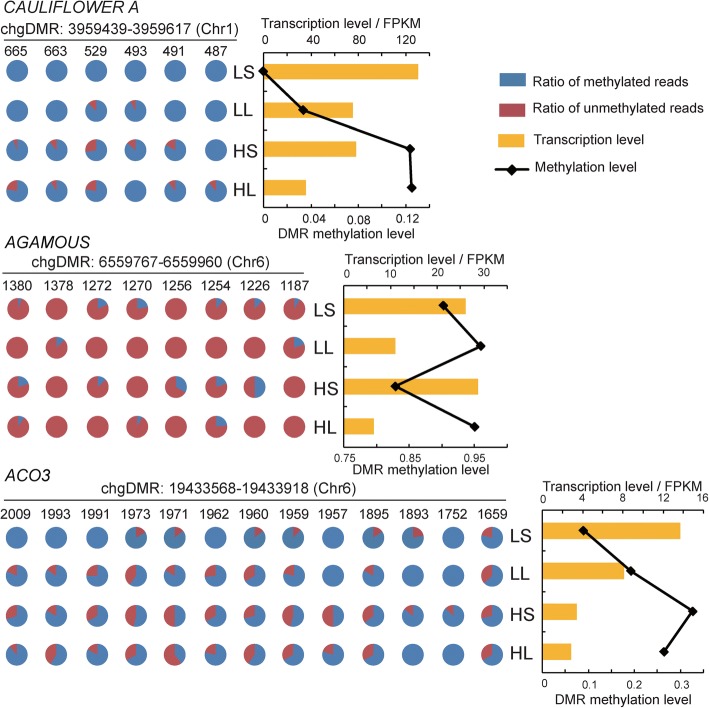
Results: The seasonal change in the femaleness of a cucumber core germplasm collection was investigated over five consecutive years. As a result, 71.3% of the 359 cucumber accessions significantly decreased their femaleness in early autumn when compared with spring. High temperature and long-day photoperiod treatments, which mimic early autumn conditions, are both unfavorable for female flower formation, and temperature is the predominant factor. High temperatures and long-day treatments both predominantly resulted in hypermethylation compared to demethylation, and temperature effect was decisive. The targeted cytosines shared in high-temperature and long-day photoperiod treatment showed the same change in DNA methylation level. Moreover, differentially expressed TEs (DETs) and the predicted epiregulation sites were clustered across chromosomes, and importantly, these sites were reproducible among different treatments. Essentially, the photoperiod treatment preferentially and significantly influenced flower development processes, while temperature treatment produced stronger responses from phytohormone-pathway-related genes. Cucumber AGAMOUS was likely epicontrolled exclusively by photoperiod while CAULIFLOWER A and CsACO3 were likely epicontrolled by both photoperiod and temperature.
Conclusions: Seasonal change of sex expression is a germplasm-wide phenomenon in cucumbers. High temperature and long-day photoperiod might have the same effect on the methylome via the same mechanism of gene-TE interaction but resulted in different epicontrol sites that account for different mechanisms between temperature- and photoperiod-dependent sex expression changes.
Keywords: Cucumber germplasm; DNA methylation; Photoperiod; Sex expression; Temperature.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Cantliffe DJ. Alteration of sex expression in cucumber due to changes in temperature. light intensity and photoperiod J Am Soc Hortic Sci. 1981;106:133–136.
-
- Bachman GR, McMahon MJ. Light quality effects on flower sex expression in cucumber and spinach. HortScience. 1997;32:464.
-
- Ito H, Saito T. Factors responsible for sex expression of Japanese cucumber. IX. Effects of nitrogen application and watering under the controlled day length and night temperature in the nursery bed. J Jpn Soc Hortic Sci. 1958;27:11–19. doi: 10.2503/jjshs.27.11. - DOI
-
- Takahashi H, Suge H. Sex expression in cucumber plants as affected by mechanical stress. Plant Cell Physiol. 1980;21:303–310. doi: 10.1093/oxfordjournals.pcp.a076003. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
