

Hypoxia Promotes Immune Evasion by Triggering β-Glucan Masking on the Candida albicans Cell Surface via Mitochondrial and cAMP-Protein Kinase A Signaling
- PMID: 30401773
- PMCID: PMC6222127
- DOI: 10.1128/mBio.01318-18
Hypoxia Promotes Immune Evasion by Triggering β-Glucan Masking on the Candida albicans Cell Surface via Mitochondrial and cAMP-Protein Kinase A Signaling
Abstract
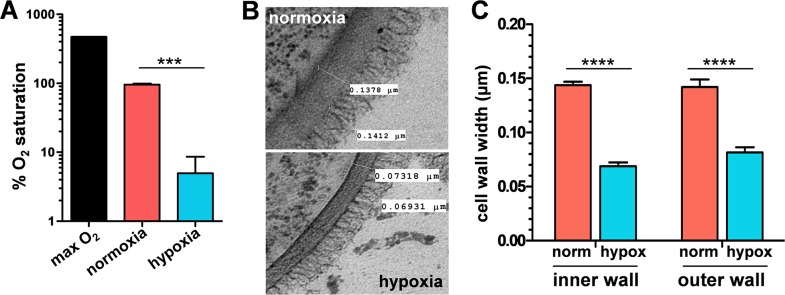
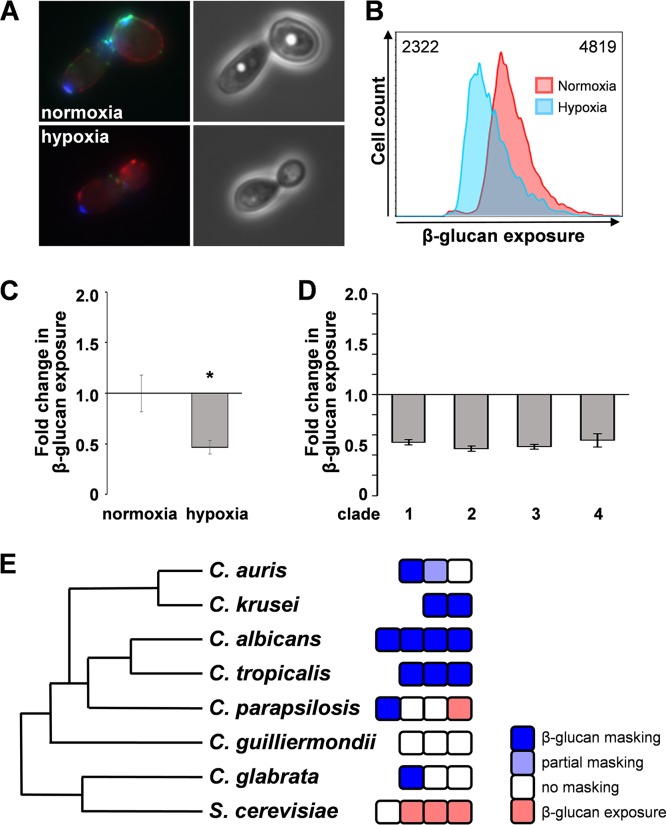
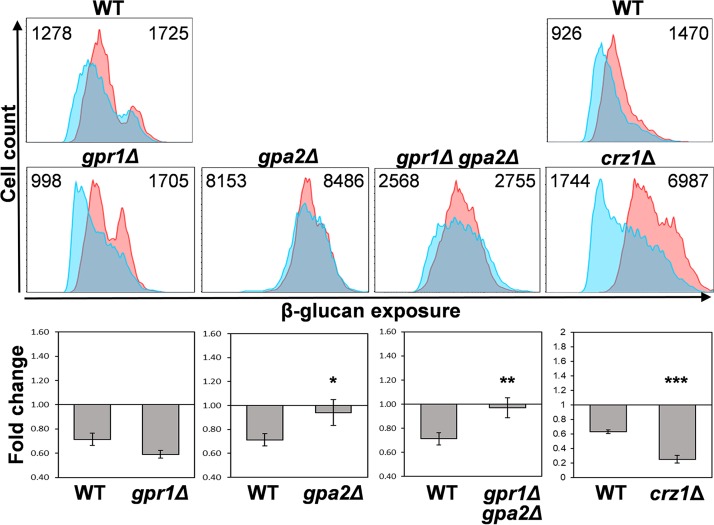
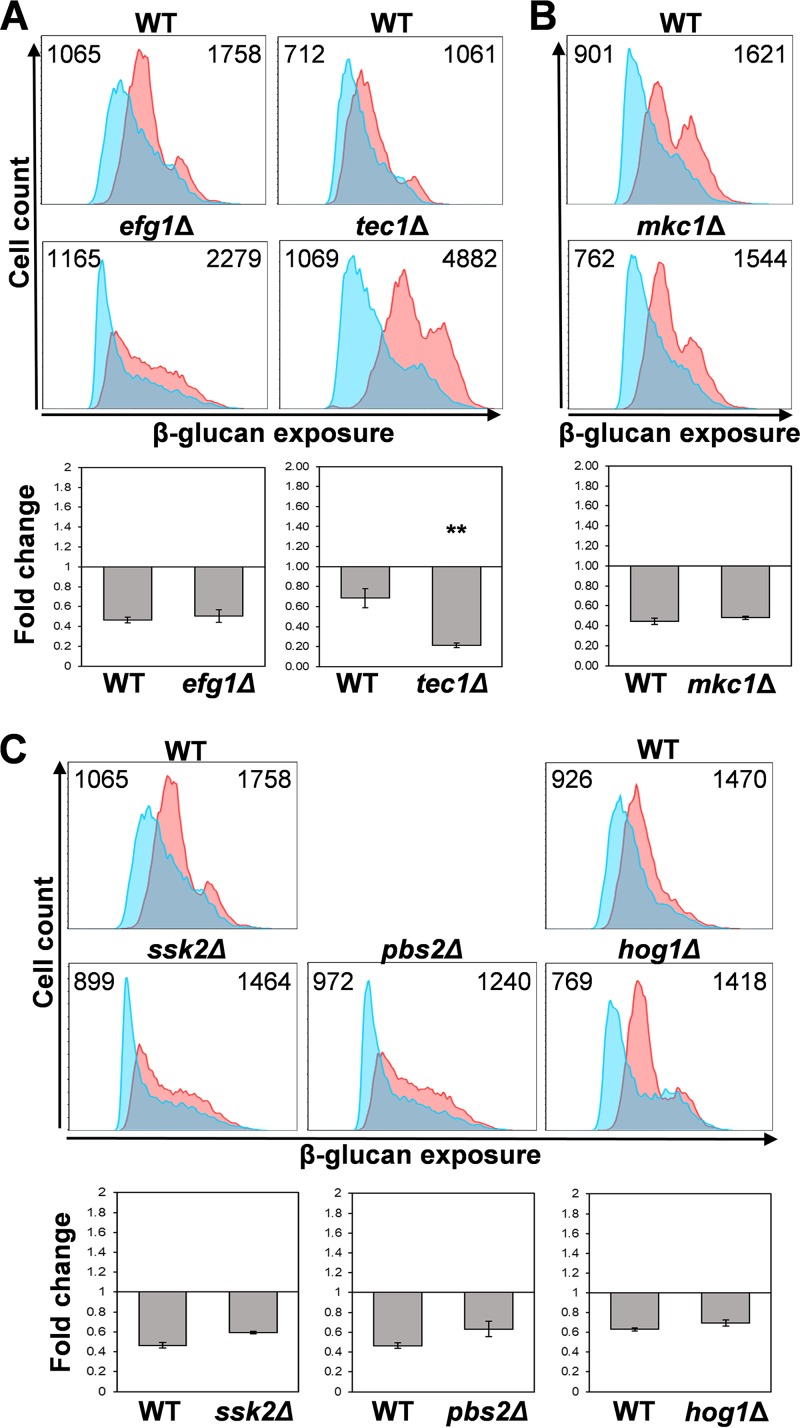
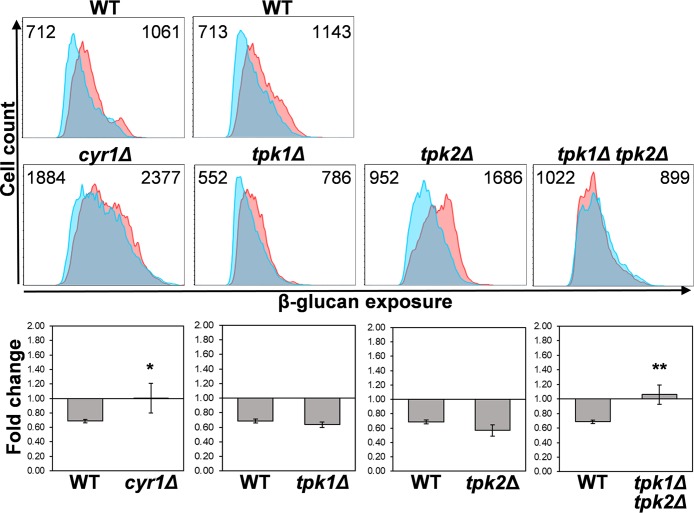
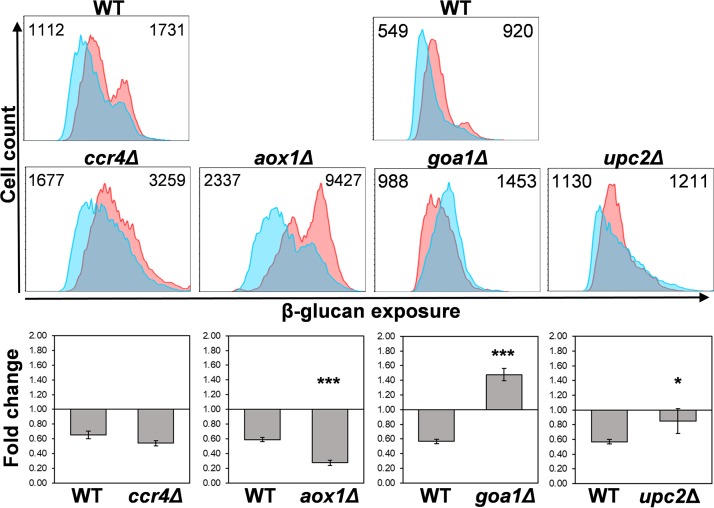
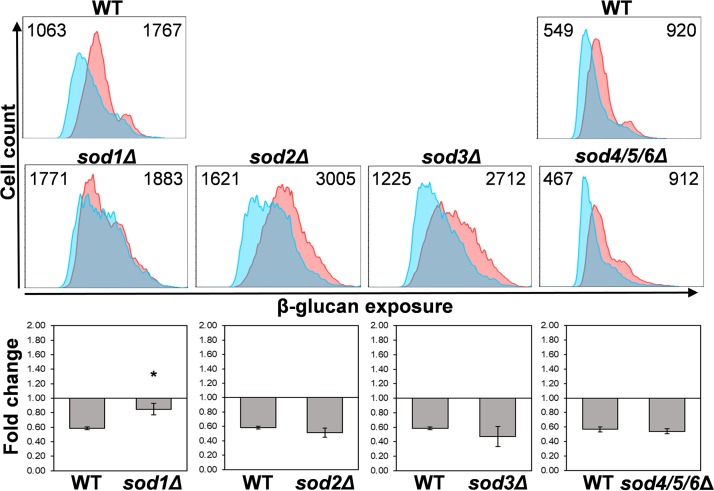
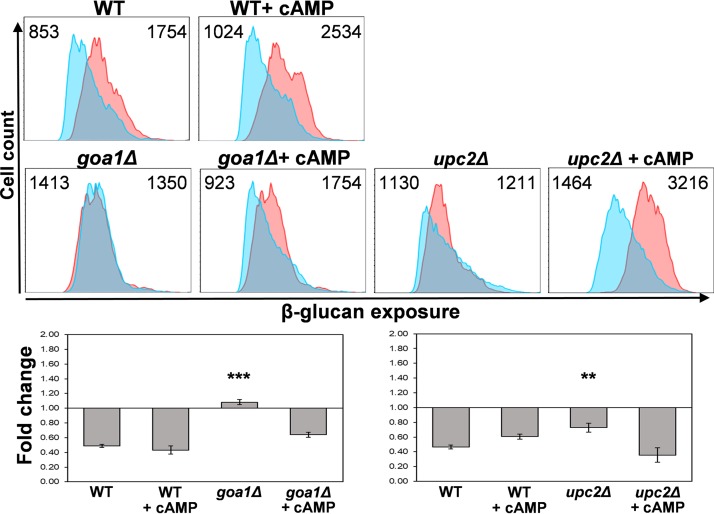
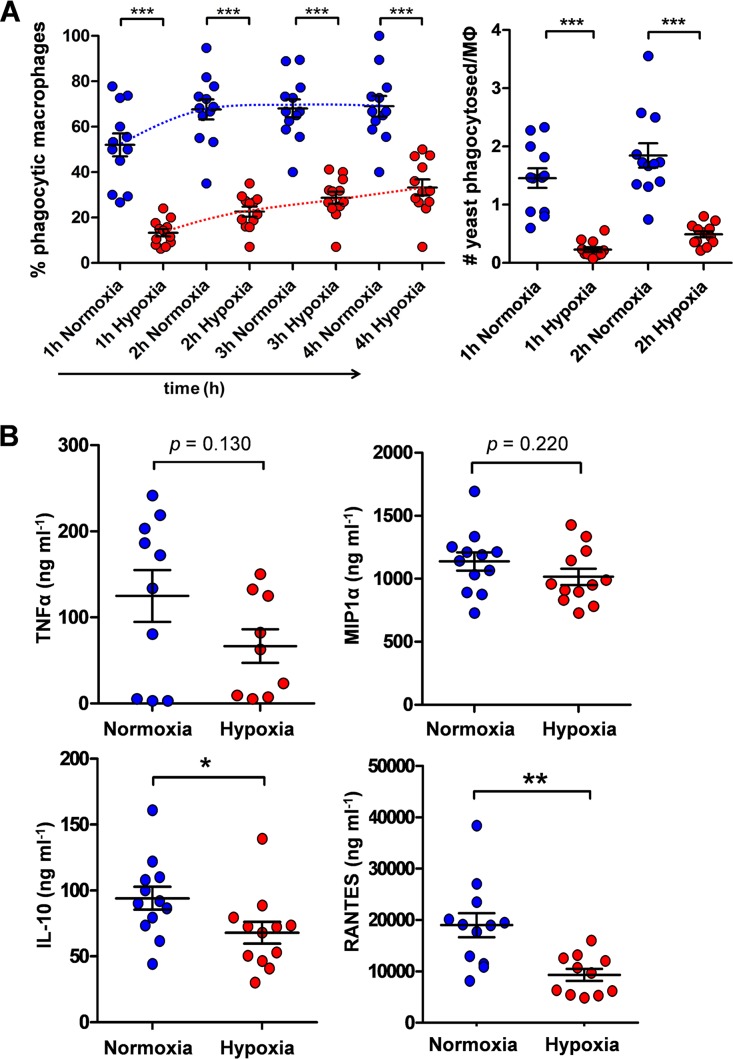
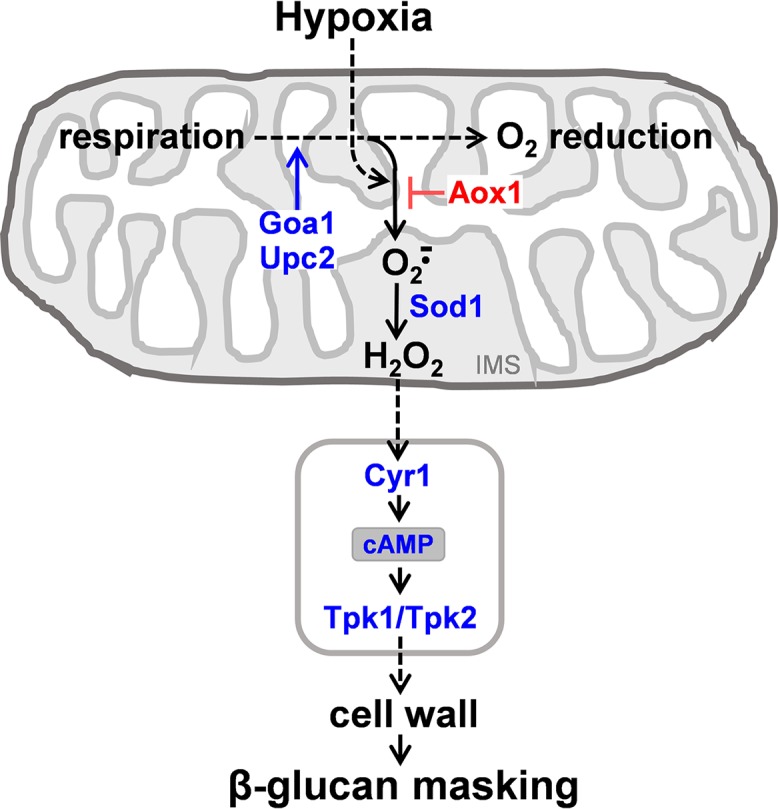
Organisms must adapt to changes in oxygen tension if they are to exploit the energetic benefits of reducing oxygen while minimizing the potentially damaging effects of oxidation. Consequently, organisms in all eukaryotic kingdoms display robust adaptation to hypoxia (low oxygen levels). This is particularly important for fungal pathogens that colonize hypoxic niches in the host. We show that adaptation to hypoxia in the major fungal pathogen of humans Candida albicans includes changes in cell wall structure and reduced exposure, at the cell surface, of β-glucan, a key pathogen-associated molecular pattern (PAMP). This leads to reduced phagocytosis by murine bone marrow-derived macrophages and decreased production of IL-10, RANTES, and TNF-α by peripheral blood mononuclear cells, suggesting that hypoxia-induced β-glucan masking has a significant effect upon C. albicans-host interactions. We show that hypoxia-induced β-glucan masking is dependent upon both mitochondrial and cAMP-protein kinase A (PKA) signaling. The decrease in β-glucan exposure is blocked by mutations that affect mitochondrial functionality (goa1Δ and upc2Δ) or that decrease production of hydrogen peroxide in the inner membrane space (sod1Δ). Furthermore, β-glucan masking is enhanced by mutations that elevate mitochondrial reactive oxygen species (aox1Δ). The β-glucan masking defects displayed by goa1Δ and upc2Δ cells are suppressed by exogenous dibutyryl-cAMP. Also, mutations that inactivate cAMP synthesis (cyr1Δ) or PKA (tpk1Δ tpk2Δ) block the masking phenotype. Our data suggest that C. albicans responds to hypoxic niches by inducing β-glucan masking via a mitochondrial cAMP-PKA signaling pathway, thereby modulating local immune responses and promoting fungal colonization.IMPORTANCE Animal, plant, and fungal cells occupy environments that impose changes in oxygen tension. Consequently, many species have evolved mechanisms that permit robust adaptation to these changes. The fungal pathogen Candida albicans can colonize hypoxic (low oxygen) niches in its human host, such as the lower gastrointestinal tract and inflamed tissues, but to colonize its host, the fungus must also evade local immune defenses. We reveal, for the first time, a defined link between hypoxic adaptation and immune evasion in C. albicans As this pathogen adapts to hypoxia, it undergoes changes in cell wall structure that include masking of β-glucan at its cell surface, and it becomes better able to evade phagocytosis by innate immune cells. We also define the signaling mechanisms that mediate hypoxia-induced β-glucan masking, showing that they are dependent on mitochondrial signaling and the cAMP-protein kinase pathway. Therefore, hypoxia appears to trigger immune evasion in this fungal pathogen.
Keywords: Candida albicans; cAMP-protein kinase A signaling; cell wall; hypoxia; mitochondrial signaling; β-glucan masking.
Copyright © 2018 Pradhan et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
