

Review
doi: 10.1155/2018/5207608.
eCollection 2018.
Exploring Seipin: From Biochemistry to Bioinformatics Predictions
Affiliations
- PMID: 30402103
- PMCID: PMC6192094
- DOI: 10.1155/2018/5207608
Item in Clipboard
Review
Exploring Seipin: From Biochemistry to Bioinformatics Predictions
Int J Cell Biol.
.
Abstract
Seipin is a nonenzymatic protein encoded by the BSCL2 gene. It is involved in lipodystrophy and seipinopathy diseases. Named in 2001, all seipin functions are still far from being understood. Therefore, we reviewed much of the research, trying to find a pattern that could explain commonly observed features of seipin expression disorders. Likewise, this review shows how this protein seems to have tissue-specific functions. In an integrative view, we conclude by proposing a theoretical model to explain how seipin might be involved in the triacylglycerol synthesis pathway.
Figures

Human seipin isoforms. Multiple alignments of seipin isoforms were performed through T-Coffee [32]. Isoform 3 is the biggest with 462 amino acids, followed by 1 and 2 with 398 and 287 amino acids, respectively. UniProt [14] considers seipin isoform 1 as the canonical one. Pink color represents identical alignments; yellow corresponds to similar alignments; and green regions show different alignments. ∗ corresponds to an equal match and the differences are highlighted by . and : symbols. Cons: consensus sequence; Iso: isoform.
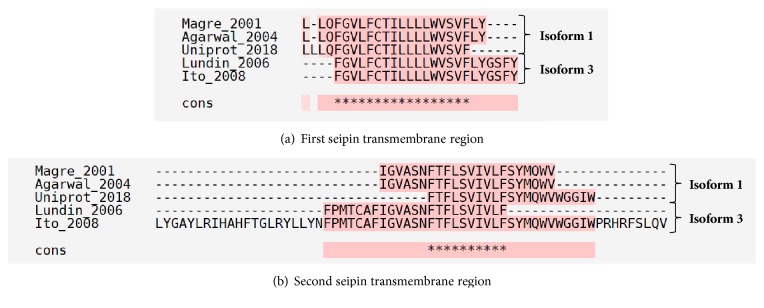
Alignment of transmembrane regions of seipin. Multiple alignments of seipin isoforms were performed through T-Coffee [32]. Many authors predicted the transmembrane regions of seipin and the amino acid positions are reviewed in Table 1 [4, 6, 14, 33, 34]. It is possible to observe that, even with differences, some regions are conserved in the prediction for the same isoform or between different isoforms. Isoform 2 was omitted due to the low number of works with it. Pink color represents identical alignments; yellow corresponds to similar alignments; and green regions show different alignments. ∗ corresponds to an equal match. Cons: consensus sequence.
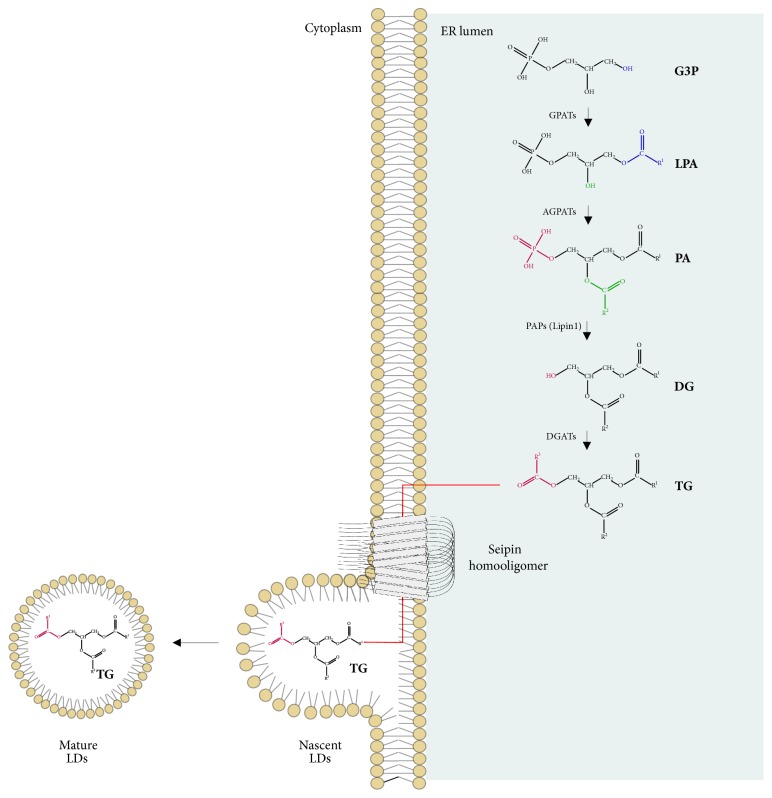
Triacylglycerol synthesis and usual seipin localization. During triacylglycerol synthesis, glycerol-3-phosphate acyltransferases (GPATs) catalyze the acylation at sn-1 position of glycerol-3-phosphate (G3P) and origin lysophosphatidic acid (LPA). Then, 1-acyl-sn-glycerol-3-phosphate acyltransferases (AGPATs) catalyze the acylation at sn-2 of LPA and give rise to phosphatidic acid (PA). Later, phosphatidate phosphatases (PAPs), as lipin1, can remove the phosphate group from PA and produce diacylglycerol (DG). Finally, diacylglycerol o-acyltransferases (DGATs) catalyze the acylation at the sn-3 position and give rise to triacylglycerol (TG) [35, 36]. In the same context, seipin comes as an oligomeric endoplasmic reticulum (ER) transmembrane protein that acts in lipid droplet (LD) assembly. ER and LDs were found to be neighbors, and seipin is concentrated in the communication regions between them, enabling the transfer of lipids recently synthetized to nascent LDs [–39]. Pieces of the illustrations are from the SMART website [40].
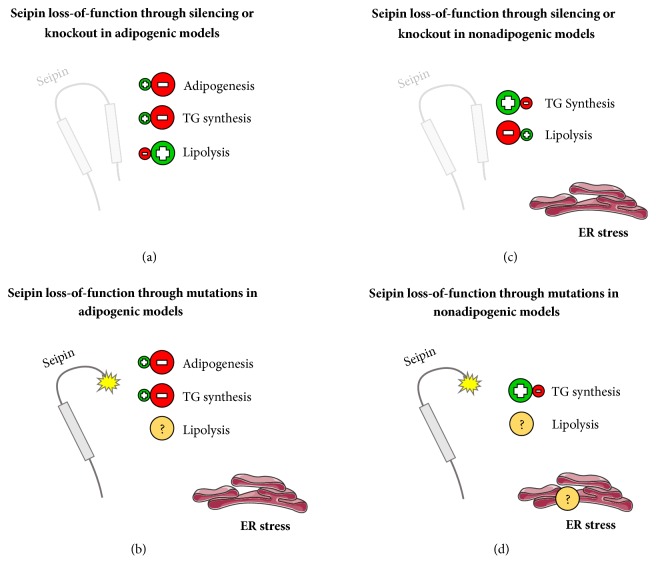
Seipin loss-of-function. We are proposing 4 different general models that usually happen with frequency under seipin loss-of-function. (a) Seipin loss-of-function through silencing or knockout in adipogenic models. Adipocyte maturation and TG synthesis are impaired, and ER stress was not found. (b) Seipin loss-of-function through mutations in adipogenic models. Adipocyte maturation and TG synthesis are impaired, ER stress was positively found, and there is a lack of information about the lipolysis situation. (c) Seipin loss-of-function through silencing or knockout in nonadipogenic models. TG synthesis was increased, lipolysis is impaired, and ER stress was positively found; (d) seipin loss-of-function through mutations in nonadipogenic models. TG synthesis was increased, and there is a lack of information about ER stress and lipolysis. (b) and (d) are the most representative situations of Berardinelli-Seip congenital lipodystrophy (BSCL) type 2. Positive symbols represent a process that is usually increased, while the negative symbols represent the opposite. The interrogation symbol represents a process that needs to be studied further. Pieces of the illustrations are from the SMART website [40].
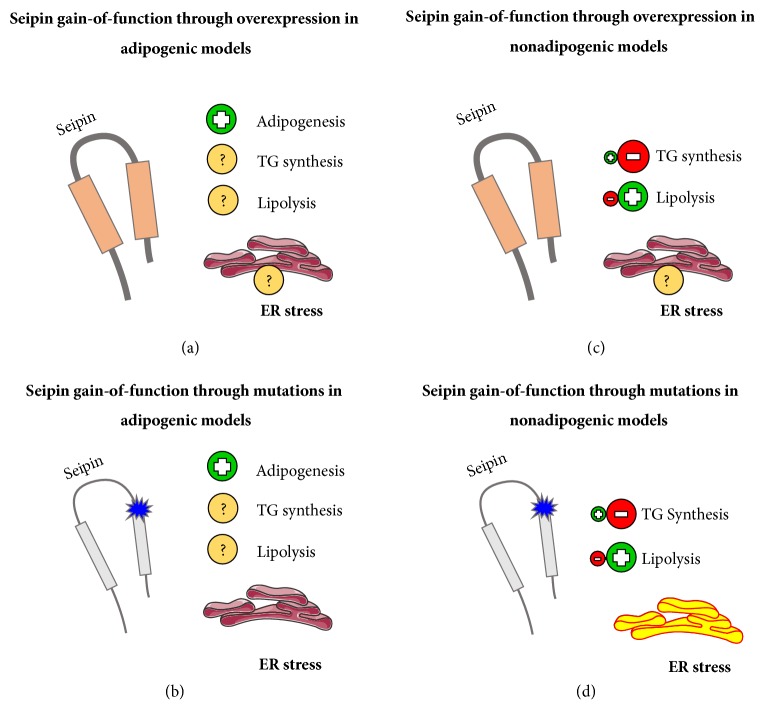
Seipin gain-of-function. We are proposing 4 different general models that usually happen with frequency under seipin gain-of-function. (a) Seipin gain-of-function through overexpression in adipogenic models. Adipocyte maturation was not impaired, but other parameters are still not clear. (b) Seipin gain-of-function through mutations in adipogenic models. Adipocyte maturation was not impaired even with ER stress. (c) Seipin gain-of-function through overexpression in nonadipogenic models. There was TG synthesis decrease, with lipolysis increase. (d) Seipin gain-of-function through mutations in nonadipogenic models. TG synthesis impairment was observed together with lipolysis increase and strong and significant ER stress. (b) and (d) models are related to seipinopathy. Positive symbols represent a process that is usually increased, while the negative symbols represent the opposite. The interrogation symbol represents a process that needs to be studied further. Pieces of the illustrations are from the SMART website [40].
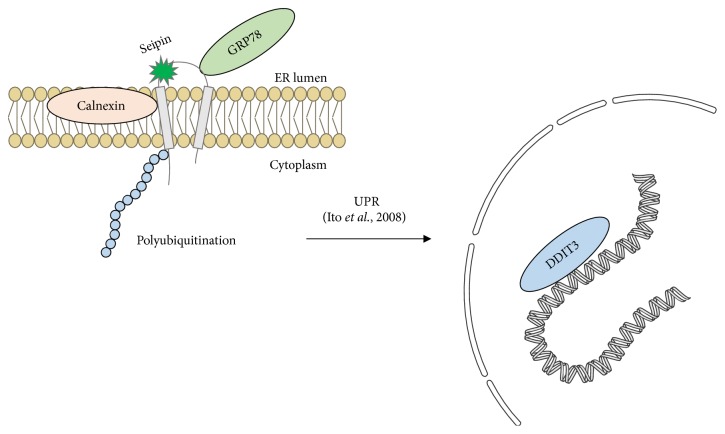
Gain-of-function mutations in seipin elicit ER stress in seipinopathies. Ito et al. proposed a model in which N88S and S90L mutations are able to disturb the seipin glycosylation site and generate ER stress and the unfolded protein response (UPR) [6]. They observed increased apoptosis as a result of the process. DDIT3 (also called CHOP) is a transcription factor responsible for the positive regulation of proapoptotic genes in response to ER stress. Calnexin (CANX) and GRP78 (also called BIP) are chaperones that work in UPR. As we reviewed, ER stress is not a phenomenon exclusively related to seipinopathy and might also be important for lipodystrophies. Pieces of the illustrations are from the SMART website [40].
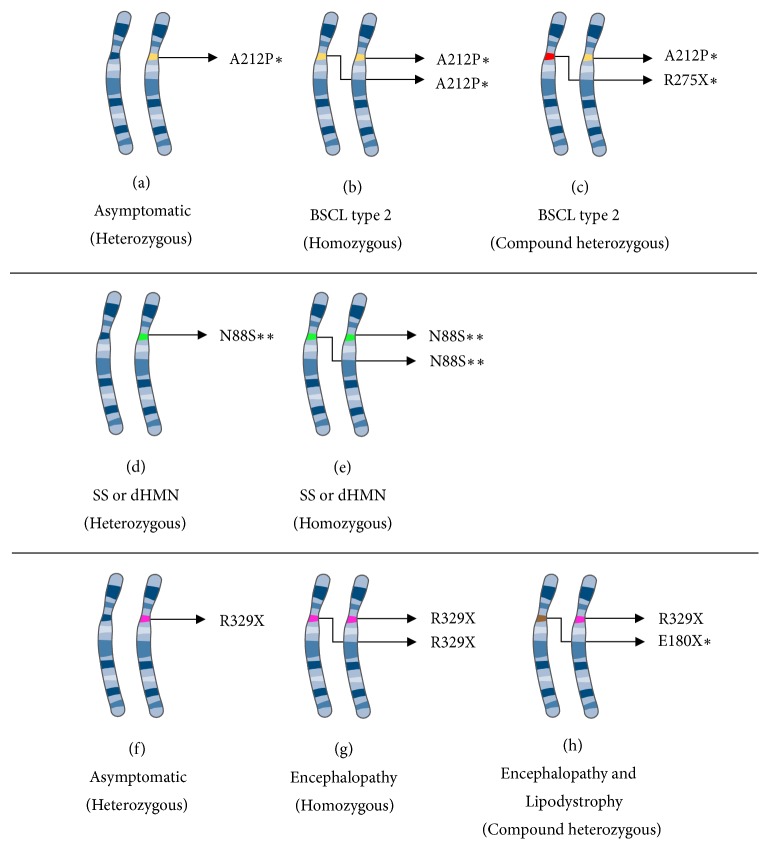
Inheritance pattern of seipin-related diseases. Here, we summarize the inheritance mechanism of seipin-related diseases. (a–c) BSCL type 2, a loss-of-function, and recessive disease. (d-e) Seipinopathies, gain-of-function, and dominant diseases. (f–h) Progressive encephalopathy with or without lipodystrophy (PELD) and a gain-of-function and recessive disease. Pieces of the illustrations are from the SMART website [40]. ∗ Or other mutations related to BSCL type 2. ∗∗ Or other mutations related to seipinopathy. BSCL: Berardinelli-Seip congenital lipodystrophy; dHMN: distal hereditary motor neuropathies; SS: Silver syndrome.
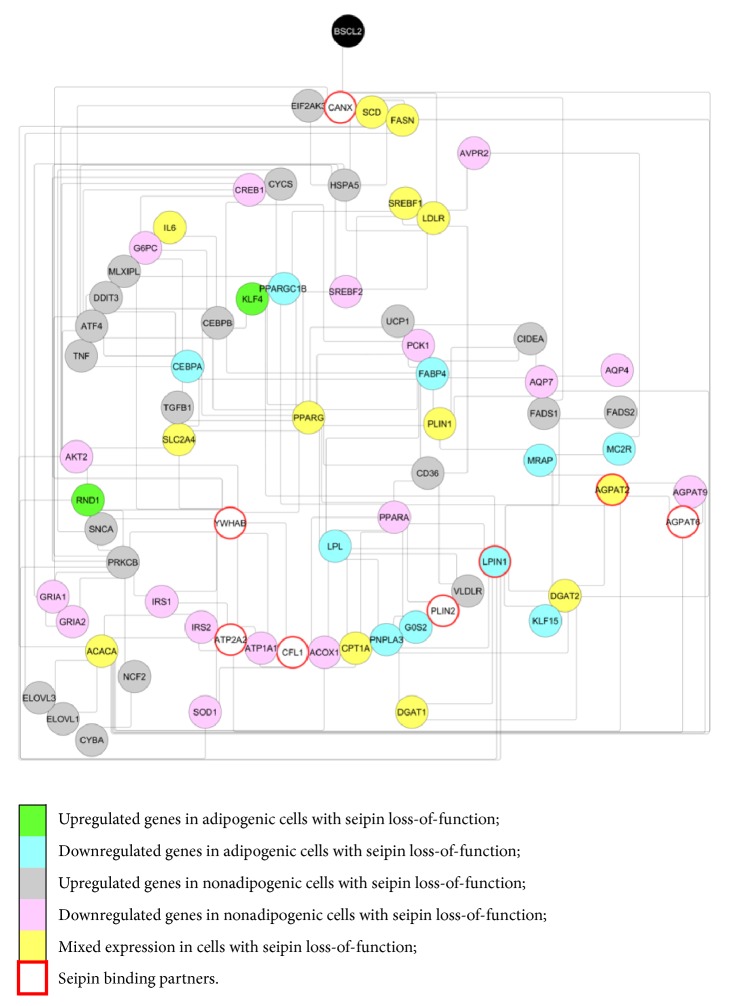
Biological integration. A network with genes differently regulated in seipin loss-of-function situations was created using STRING [41]. The gene list submitted is in accordance with Supplementary Tables 1 and 2. Genes painted with green are upregulated in adipogenic cells with seipin loss-of-function. Genes painted with blue are downregulated in adipogenic cells with seipin loss-of-function. Genes painted in gray are upregulated in nonadipogenic cells with seipin loss-of-function and genes painted in pink are downregulated in nonadipogenic cells with seipin loss-of-function. Genes that were observed either downregulated or upregulated in the same cell situations but in different papers are painted in yellow (mixed expression). Proteins already described as seipin physical binders are surrounded by a red circumference. The parameters chosen were “Experiments,” “Databases,” “Coexpression,” “Neighborhood,” “Gene Fusion,” “Cooccurrence,” and “Minimum required interaction score = 0.1.” Cytoscape [42] classifies the genes based on their connectivity. As the circle comes to the center, the node tends to be more connected with the network.
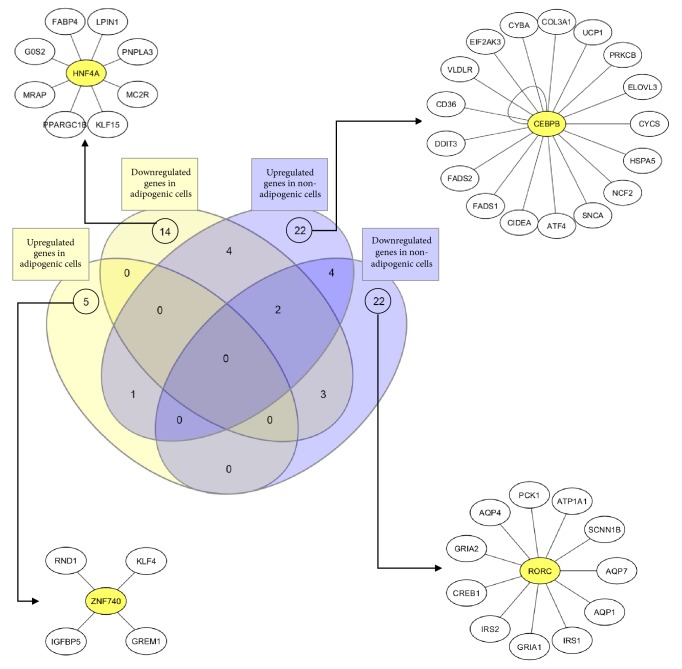
Differently expressed genes during seipin loss-of-function. InteractVenn [43] allowed us to see four different clusters formed when seipin loses its function in different cell types. iRegulon [44] shows HNF4A, CEBPB, ZNF740, and RORC as important transcription factors that may interfere with the expression of the observed genes for each cluster. Among them, only CEBPB was studied in a loss of seipin function [45].
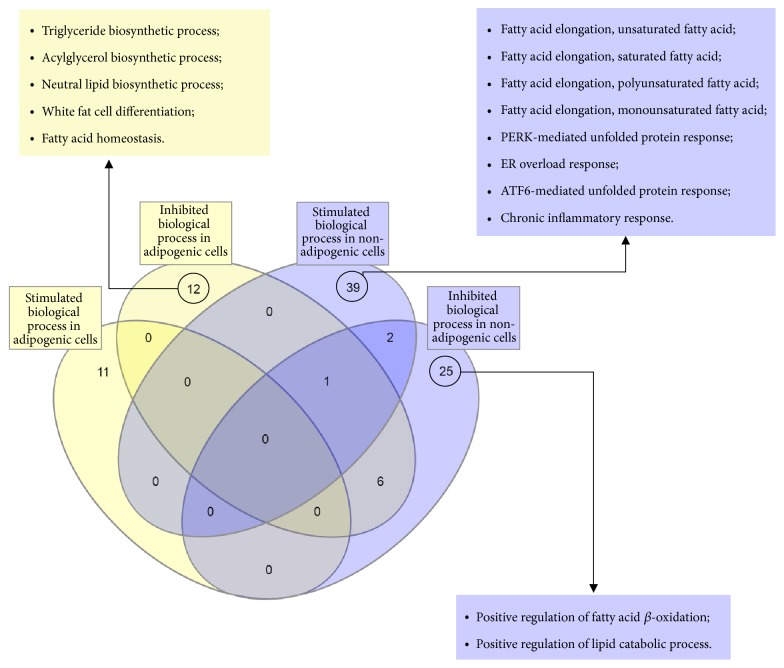
Gene ontology for seipin loss-of-function. InteractVenn [43] allowed us to see four different clusters formed when seipin loses its function in different cell types. All of the biological processes had a fold enrichment >100.
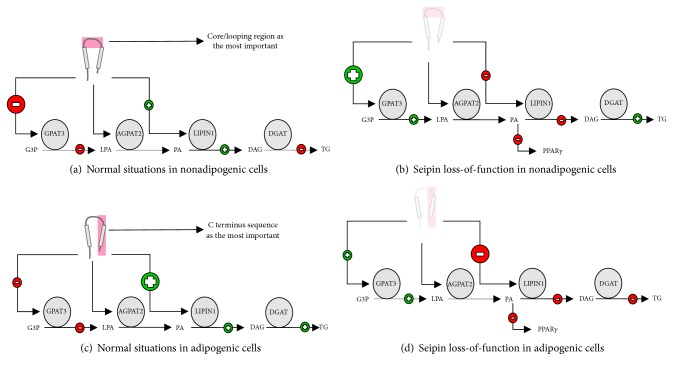
Theoretical model for seipin. (a) Seipin seems to have a core/looping region with more importance for nonadipogenic cells. This region can interact with GPAT3 and downregulate its activity, compromising TG synthesis. (b) Seipin loss-of-function in nonadipogenic cells seems to have the opposite behavior of the situation proposed in “(a)”. (c) Seipin also seems to have a C terminus region with more importance for adipogenesis. This region can interact with lipin1 and 14-3-3β to promote TG synthesis and adipogenesis. (d) Seipin loss-of-function in adipogenic cells seems to have the opposite behavior of the situation proposed in “(c)”. AGPAT: 1-acyl-sn-glycerol-3-phosphate acyltransferase; DG: diacylglycerol; DGAT: diacylglycerol o-acyltransferases; G3P: glycerol-3-phosphate; GPAT: glycerol-3-phosphate acyltransferase; LPA: lysophosphatidic acid; PA: phosphatidic acid; PPARγ: peroxisome proliferator-activated receptor gamma; TG: triacylglycerol.
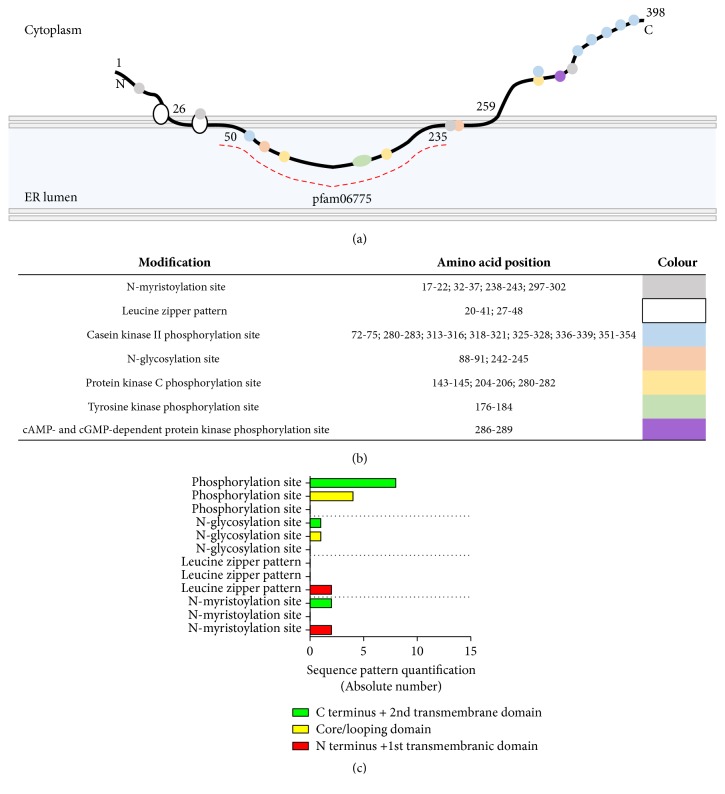
Theoretical
posttranslational modifications
of seipin. (a) Prosite [46] and Pfam [47] predictions for posttranslational modifications and conserved domains of seipin, respectively. (b) Residue patterns highlighted in (a). (c) Quantification of patterns. Even as theoretical predictions, the N-glycosylation site was already proven to occur and to be affected by the mutations N88S and S90L [48].
Similar articles
-
Function of seipin: new insights from Bscl2/seipin knockout mouse models.Biochimie. 2014 Jan;96:166-72. doi: 10.1016/j.biochi.2013.06.022. Epub 2013 Jul 2. Biochimie. 2014. PMID: 23831461 Review.
-
Overexpression of a short human seipin/BSCL2 isoform in mouse adipose tissue results in mild lipodystrophy.Am J Physiol Endocrinol Metab. 2012 Mar 15;302(6):E705-13. doi: 10.1152/ajpendo.00237.2011. Epub 2012 Jan 10. Am J Physiol Endocrinol Metab. 2012. PMID: 22234369
-
[Seipin/BSCL2-related motor neuron disease: Seipinopathy is a novel conformational disease associated with endoplasmic reticulum stress].Rinsho Shinkeigaku. 2007 Jun;47(6):329-35. Rinsho Shinkeigaku. 2007. PMID: 17633104 Review. Japanese.
-
Alleviation of seipinopathy-related ER stress by triglyceride storage.Hum Mol Genet. 2013 Mar 15;22(6):1157-66. doi: 10.1093/hmg/dds523. Epub 2012 Dec 18. Hum Mol Genet. 2013. PMID: 23250914
-
The expression of SEIPIN in the mouse central nervous system.Brain Struct Funct. 2016 Nov;221(8):4111-4127. doi: 10.1007/s00429-015-1151-3. Epub 2015 Nov 30. Brain Struct Funct. 2016. PMID: 26621102
Cited by
-
Discovering metabolic disease gene interactions by correlated effects on cellular morphology.Mol Metab. 2019 Jun;24:108-119. doi: 10.1016/j.molmet.2019.03.001. Epub 2019 Mar 13. Mol Metab. 2019. PMID: 30940487 Free PMC article.
-
Oligomers of the lipodystrophy protein seipin may co-ordinate GPAT3 and AGPAT2 enzymes to facilitate adipocyte differentiation.Sci Rep. 2020 Feb 24;10(1):3259. doi: 10.1038/s41598-020-59982-5. Sci Rep. 2020. PMID: 32094408 Free PMC article.
-
Seipin overexpression attenuates cerebral ischemia-reperfusion injury via preventing apoptosis and autophagy.Brain Behav. 2023 Dec;13(12):e3195. doi: 10.1002/brb3.3195. Epub 2023 Oct 27. Brain Behav. 2023. PMID: 37897134 Free PMC article.
-
Impaired mRNA splicing and proteostasis in preadipocytes in obesity-related metabolic disease.Elife. 2021 Sep 21;10:e65996. doi: 10.7554/eLife.65996. Elife. 2021. PMID: 34545810 Free PMC article.
-
SEIPIN: A Key Factor for Nuclear Lipid Droplet Generation and Lipid Homeostasis.Int J Mol Sci. 2020 Nov 2;21(21):8208. doi: 10.3390/ijms21218208. Int J Mol Sci. 2020. PMID: 33147895 Free PMC article. Review.
References
-
- Lima J. G., Nobrega L. H., de Lima N. N., do Nascimento Santos M. G., Baracho M. F., Jeronimo S. M. Clinical and laboratory data of a large series of patients with congenital generalized lipodystrophy. Diabetology & Metabolic Syndrome. 2016;8, article 23 doi: 10.1186/s13098-016-0140-x. - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
