

Periodicity, repression, and the molecular architecture of the mammalian circadian clock
- PMID: 30402960
- PMCID: PMC6502704
- DOI: 10.1111/ejn.14254
Periodicity, repression, and the molecular architecture of the mammalian circadian clock
Abstract
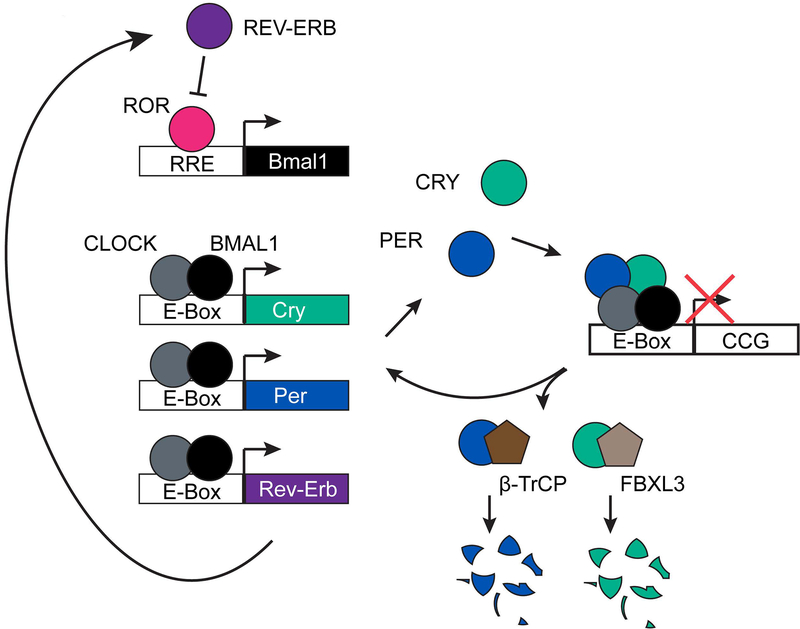
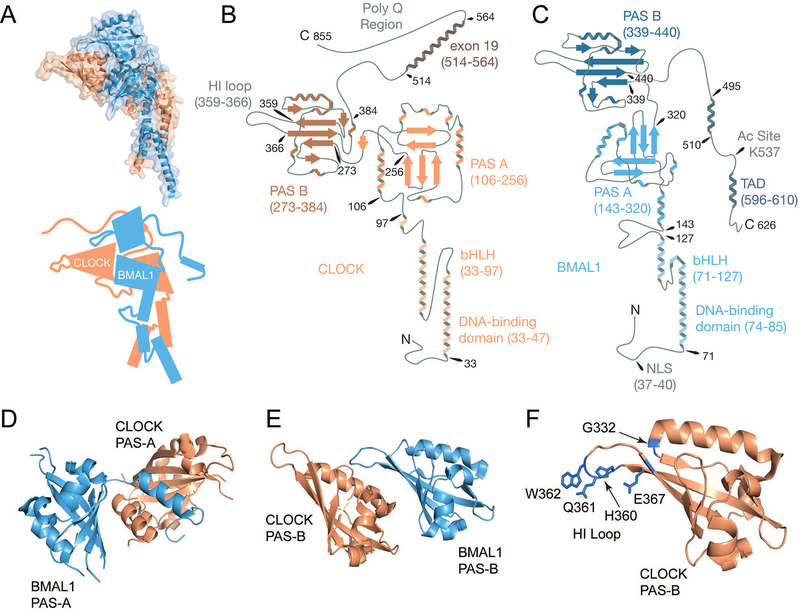
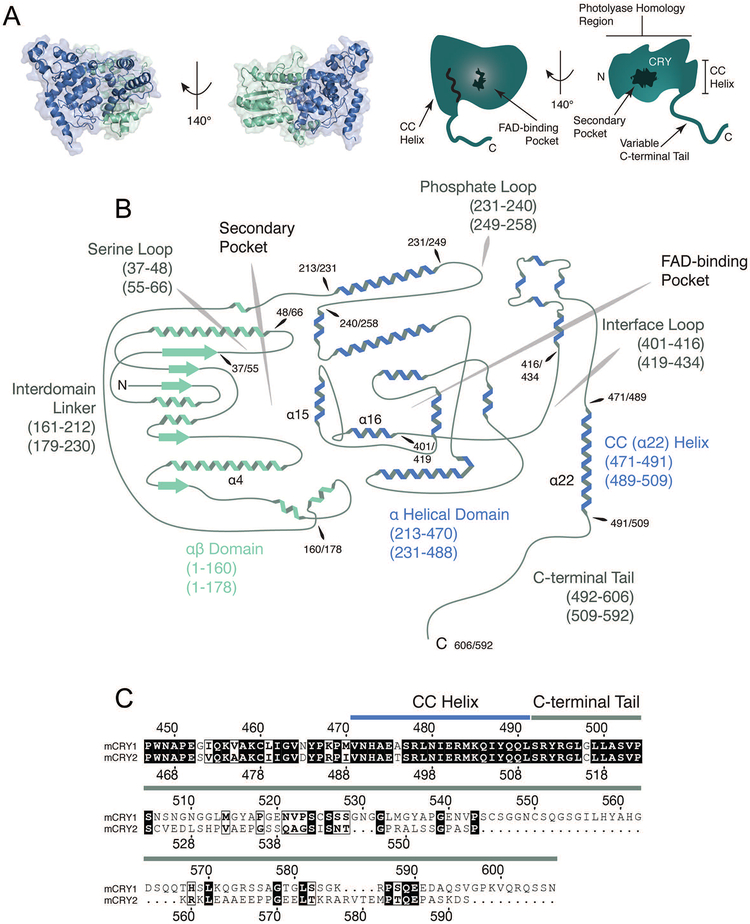
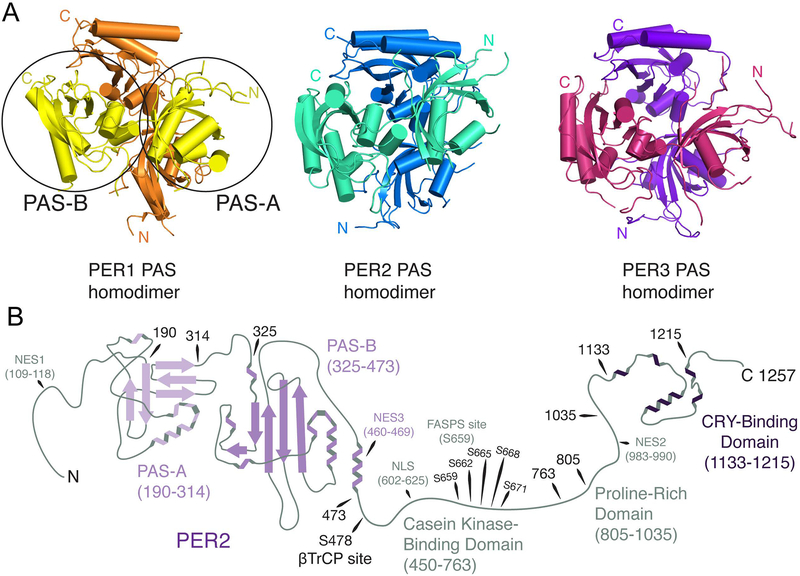
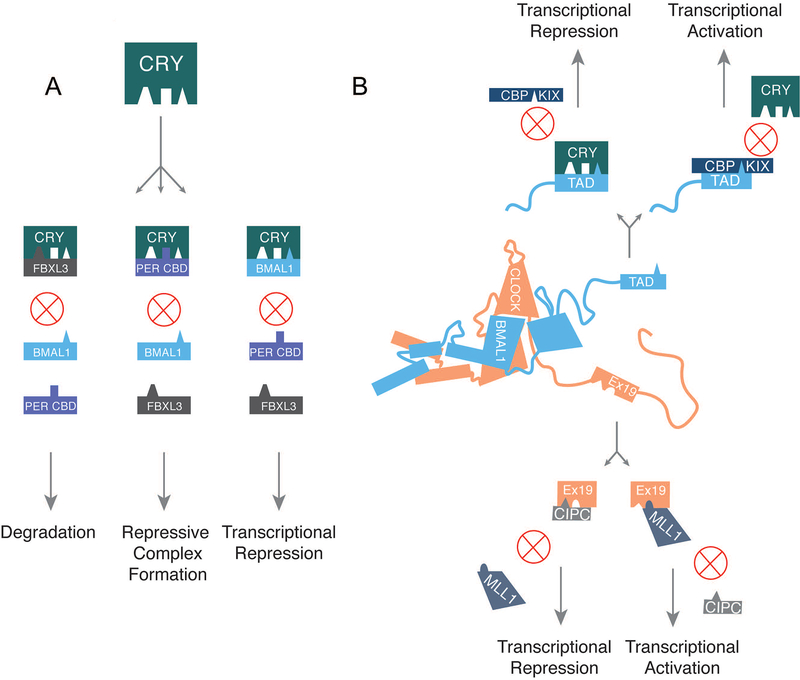
Large molecular machines regulate daily cycles of transcriptional activity and help generate rhythmic behavior. In recent years, structural and biochemical analyses have elucidated a number of principles guiding the interactions of proteins that form the basis of circadian timing. In its simplest form, the circadian clock is composed of a transcription/translation feedback loop. However, this description elides a complicated process of activator recruitment, chromatin decompaction, recruitment of coactivators, expression of repressors, formation of a repressive complex, repression of the activators, and ultimately degradation of the repressors and reinitiation of the cycle. Understanding the core principles underlying the clock requires careful examination of molecular and even atomic level details of these processes. Here, we review major structural and biochemical findings in circadian biology and make the argument that shared protein interfaces within the clockwork are critical for both the generation of rhythmicity and timing of the clock.
Keywords: clock; cryptochrome; period; rhythm; transcription.
© 2018 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Conflict of interest statement
Statement of Competing Financial Interests
All authors declare no competing financial or other interests.
Figures
Similar articles
-
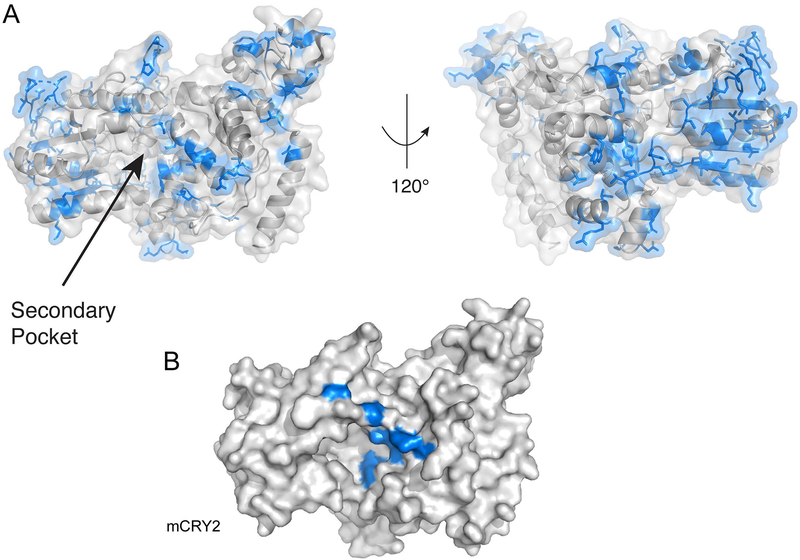
Formation of a repressive complex in the mammalian circadian clock is mediated by the secondary pocket of CRY1.Proc Natl Acad Sci U S A. 2017 Feb 14;114(7):1560-1565. doi: 10.1073/pnas.1615310114. Epub 2017 Jan 31. Proc Natl Acad Sci U S A. 2017. PMID: 28143926 Free PMC article.
-
Cycling of CRYPTOCHROME proteins is not necessary for circadian-clock function in mammalian fibroblasts.Curr Biol. 2007 Jul 3;17(13):1091-100. doi: 10.1016/j.cub.2007.05.048. Epub 2007 Jun 21. Curr Biol. 2007. PMID: 17583506 Free PMC article.
-
The circadian clock, aging and its implications in cancer.Neoplasia. 2023 Jul;41:100904. doi: 10.1016/j.neo.2023.100904. Epub 2023 May 4. Neoplasia. 2023. PMID: 37148656 Free PMC article.
-
Molecular analysis of mammalian circadian rhythms.Annu Rev Physiol. 2001;63:647-76. doi: 10.1146/annurev.physiol.63.1.647. Annu Rev Physiol. 2001. PMID: 11181971 Review.
-
Circadian oscillator proteins across the kingdoms of life: structural aspects.BMC Biol. 2019 Feb 18;17(1):13. doi: 10.1186/s12915-018-0623-3. BMC Biol. 2019. PMID: 30777051 Free PMC article. Review.
Cited by
-
The duper mutation reveals previously unsuspected functions of Cryptochrome 1 in circadian entrainment and heart disease.Proc Natl Acad Sci U S A. 2022 Aug 9;119(32):e2121883119. doi: 10.1073/pnas.2121883119. Epub 2022 Aug 5. Proc Natl Acad Sci U S A. 2022. PMID: 35930669 Free PMC article.
-
Evolution of canonical circadian clock genes underlies unique sleep strategies of marine mammals for secondary aquatic adaptation.PLoS Genet. 2025 Mar 18;21(3):e1011598. doi: 10.1371/journal.pgen.1011598. eCollection 2025 Mar. PLoS Genet. 2025. PMID: 40101169 Free PMC article.
-
The Arg-293 of Cryptochrome1 is responsible for the allosteric regulation of CLOCK-CRY1 binding in circadian rhythm.J Biol Chem. 2020 Dec 11;295(50):17187-17199. doi: 10.1074/jbc.RA120.014333. Epub 2020 Oct 7. J Biol Chem. 2020. PMID: 33028638 Free PMC article.
-
Timing Matters: The Interplay between Early Mealtime, Circadian Rhythms, Gene Expression, Circadian Hormones, and Metabolism-A Narrative Review.Clocks Sleep. 2023 Sep 6;5(3):507-535. doi: 10.3390/clockssleep5030034. Clocks Sleep. 2023. PMID: 37754352 Free PMC article. Review.
-
Phosphorylation of DNA-binding domains of CLOCK-BMAL1 complex for PER-dependent inhibition in circadian clock of mammalian cells.Proc Natl Acad Sci U S A. 2024 Jun 4;121(23):e2316858121. doi: 10.1073/pnas.2316858121. Epub 2024 May 28. Proc Natl Acad Sci U S A. 2024. PMID: 38805270 Free PMC article.
References
-
- Akashi M, Okamoto A, Tsuchiya Y, Todo T, Nishida E, and Node K (2014). A positive role for PERIOD in mammalian circadian gene expression. Cell reports 7, 1056–1064. - PubMed
-
- Albrecht U, Sun ZS, Eichele G, and Lee CC (1997). A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell 91, 1055–1064. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
