

Evidence that regulation of intramembrane proteolysis is mediated by substrate gating during sporulation in Bacillus subtilis
- PMID: 30403663
- PMCID: PMC6242693
- DOI: 10.1371/journal.pgen.1007753
Evidence that regulation of intramembrane proteolysis is mediated by substrate gating during sporulation in Bacillus subtilis
Abstract
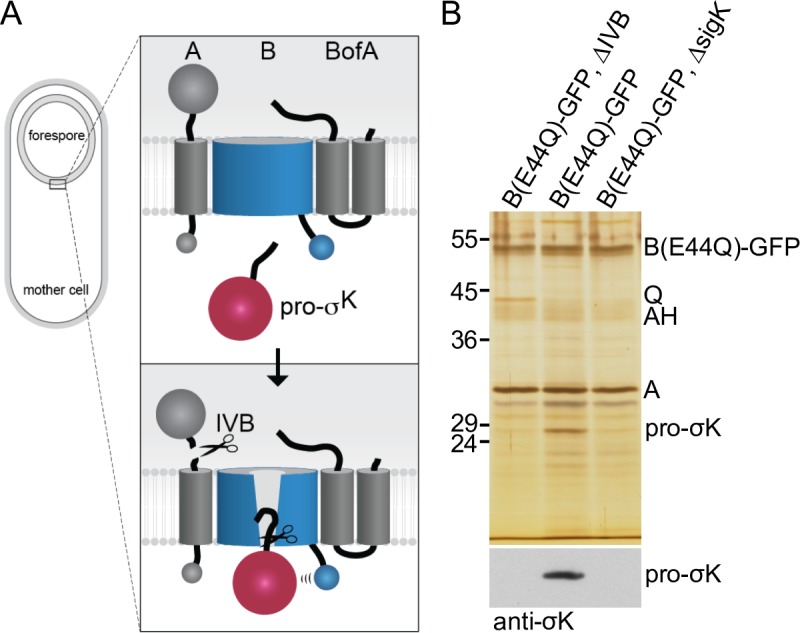
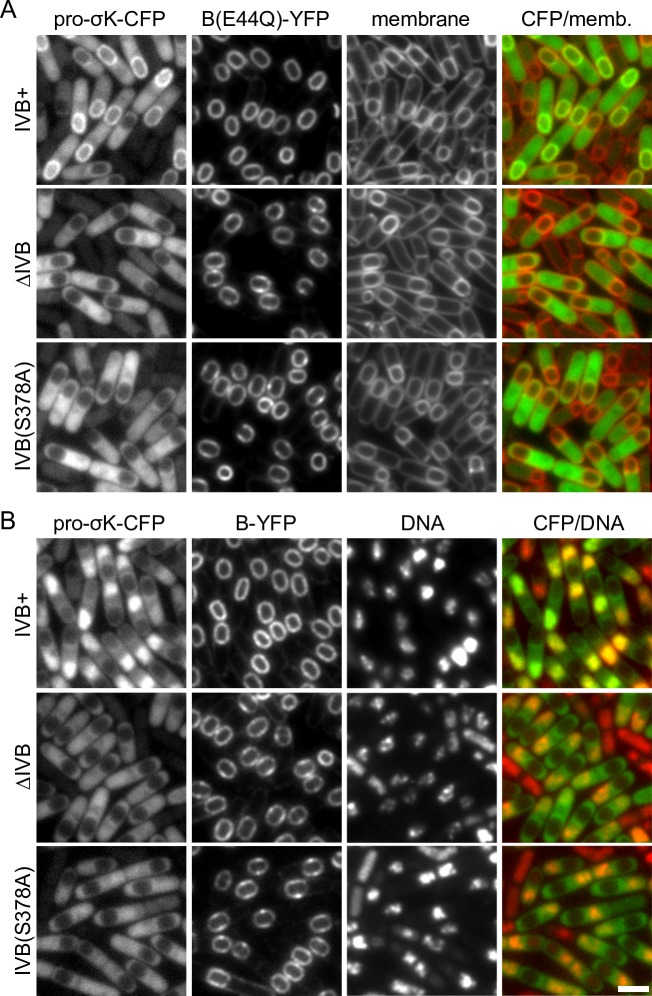
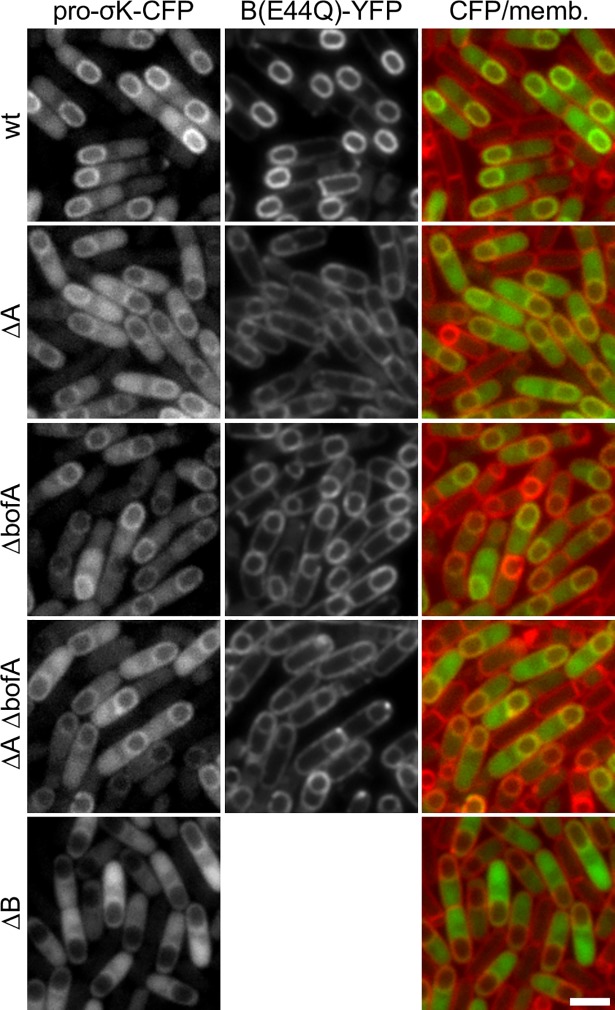
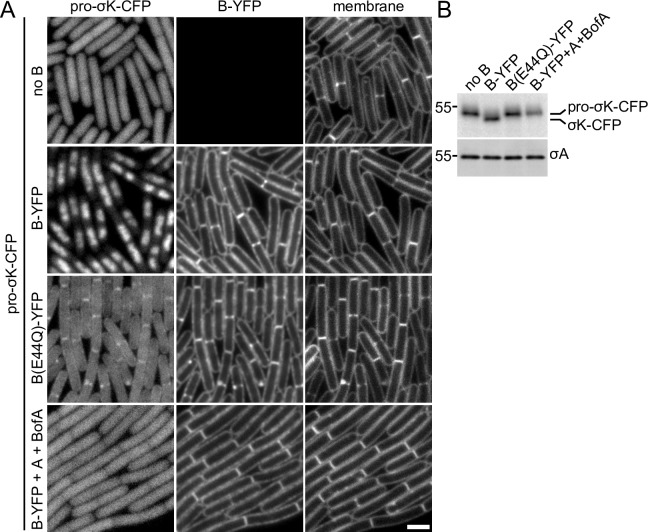
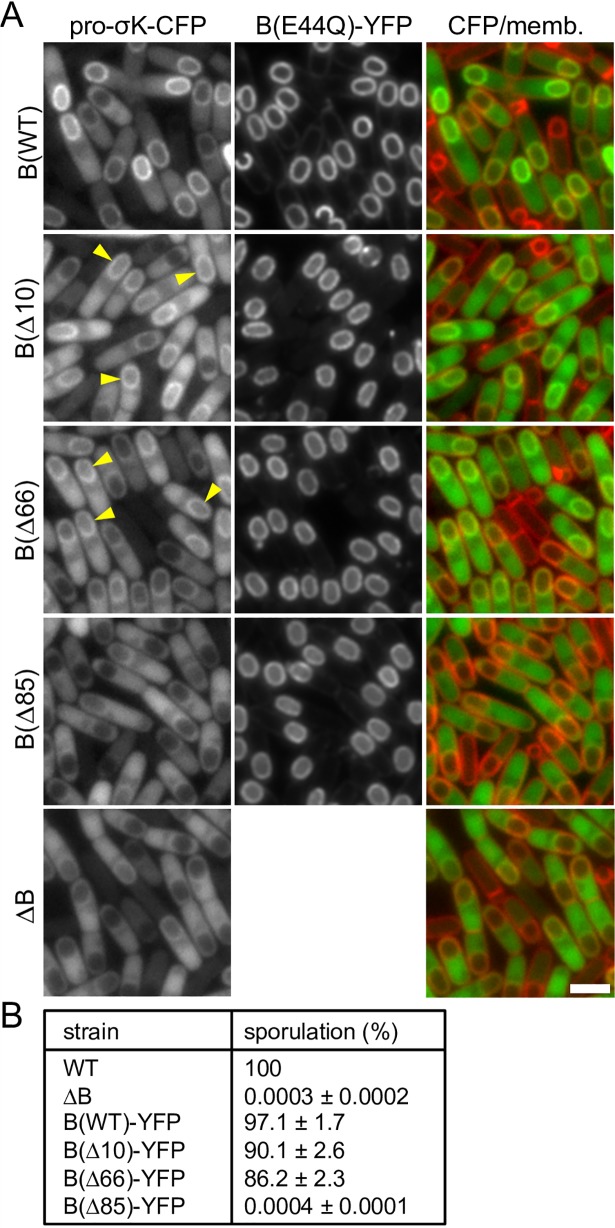
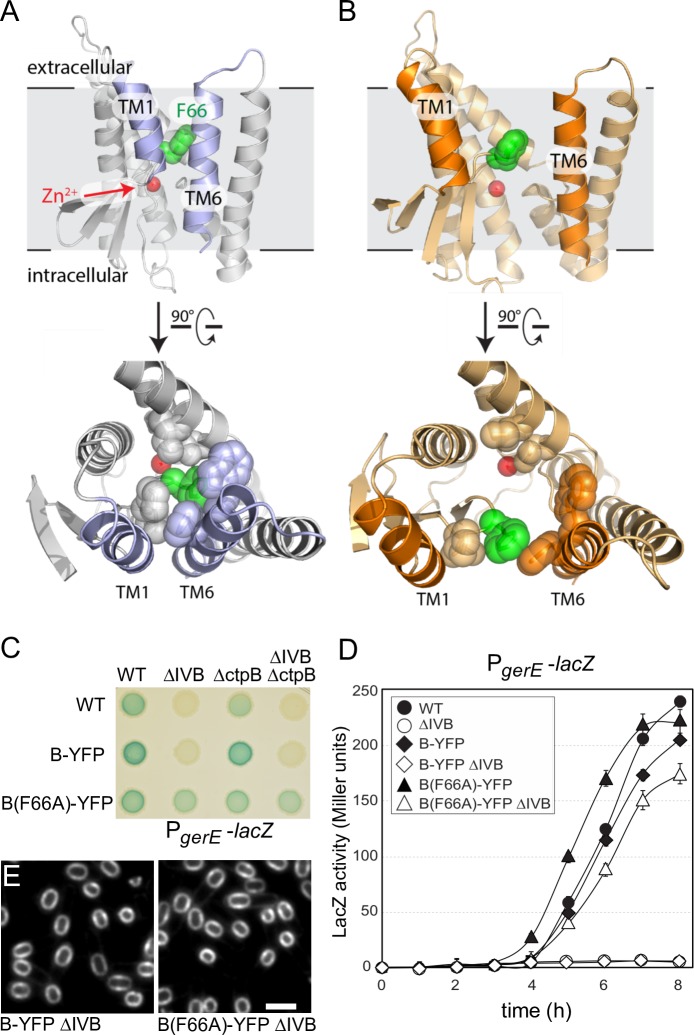
During the morphological process of sporulation in Bacillus subtilis two adjacent daughter cells (called the mother cell and forespore) follow different programs of gene expression that are linked to each other by signal transduction pathways. At a late stage in development, a signaling pathway emanating from the forespore triggers the proteolytic activation of the mother cell transcription factor σK. Cleavage of pro-σK to its mature and active form is catalyzed by the intramembrane cleaving metalloprotease SpoIVFB (B), a Site-2 Protease (S2P) family member. B is held inactive by two mother-cell membrane proteins SpoIVFA (A) and BofA. Activation of pro-σK processing requires a site-1 signaling protease SpoIVB (IVB) that is secreted from the forespore into the space between the two cells. IVB cleaves the extracellular domain of A but how this cleavage activates intramembrane proteolysis has remained unclear. Structural studies of the Methanocaldococcus jannaschii S2P homolog identified closed (substrate-occluded) and open (substrate-accessible) conformations of the protease, but the biological relevance of these conformations has not been established. Here, using co-immunoprecipitation and fluorescence microscopy, we show that stable association between the membrane-embedded protease and its substrate requires IVB signaling. We further show that the cytoplasmic cystathionine-β-synthase (CBS) domain of the B protease is not critical for this interaction or for pro-σK processing, suggesting the IVB-dependent interaction site is in the membrane protease domain. Finally, we provide evidence that the B protease domain adopts both open and closed conformations in vivo. Collectively, our data support a substrate-gating model in which IVB-dependent cleavage of A on one side of the membrane triggers a conformational change in the membrane-embedded protease from a closed to an open state allowing pro-σK access to the caged interior of the protease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Brown MS, Ye J, Rawson RB, Goldstein JL. Regulated intramembrane proteolysis: a control mechanism conserved from bacteria to humans. Cell. 2000;100(4):391–8. . - PubMed
-
- Sun L, Li X, Shi Y. Structural biology of intramembrane proteases: mechanistic insights from rhomboid and S2P to gamma-secretase. Curr Opin Struct Biol. 2016;37:97–107. 10.1016/j.sbi.2015.12.008 . - DOI - PubMed
-
- Wolfe MS, Kopan R. Intramembrane proteolysis: theme and variations. Science. 2004;305(5687):1119–23. 10.1126/science.1096187 . - DOI - PubMed
-
- Kroos L, Akiyama Y. Biochemical and structural insights into intramembrane metalloprotease mechanisms. Biochim Biophys Acta. 2013;1828(12):2873–85. 10.1016/j.bbamem.2013.03.032 ; PubMed Central PMCID: PMCPMC3793210. - DOI - PMC - PubMed
-
- Rawson RB. The site-2 protease. Biochim Biophys Acta. 2013;1828(12):2801–7. 10.1016/j.bbamem.2013.03.031 . - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
