

Evolution and functional characterization of pectate lyase PEL12, a member of a highly expanded Clonostachys rosea polysaccharide lyase 1 family
- PMID: 30404596
- PMCID: PMC6223089
- DOI: 10.1186/s12866-018-1310-9
Evolution and functional characterization of pectate lyase PEL12, a member of a highly expanded Clonostachys rosea polysaccharide lyase 1 family
Abstract
Background: Pectin is one of the major and most complex plant cell wall components that needs to be overcome by microorganisms as part of their strategies for plant invasion or nutrition. Microbial pectinolytic enzymes therefore play a significant role for plant-associated microorganisms and for the decomposition and recycling of plant organic matter. Recently, comparative studies revealed significant gene copy number expansion of the polysaccharide lyase 1 (PL1) pectin/pectate lyase gene family in the Clonostachys rosea genome, while only low numbers were found in Trichoderma species. Both of these fungal genera are widely known for their ability to parasitize and kill other fungi (mycoparasitism) and certain species are thus used for biocontrol of plant pathogenic fungi.
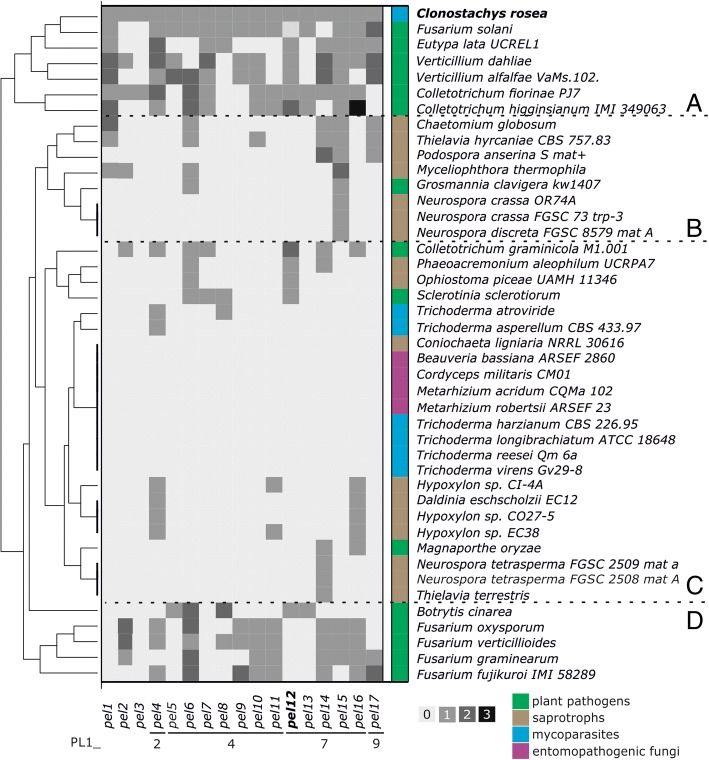
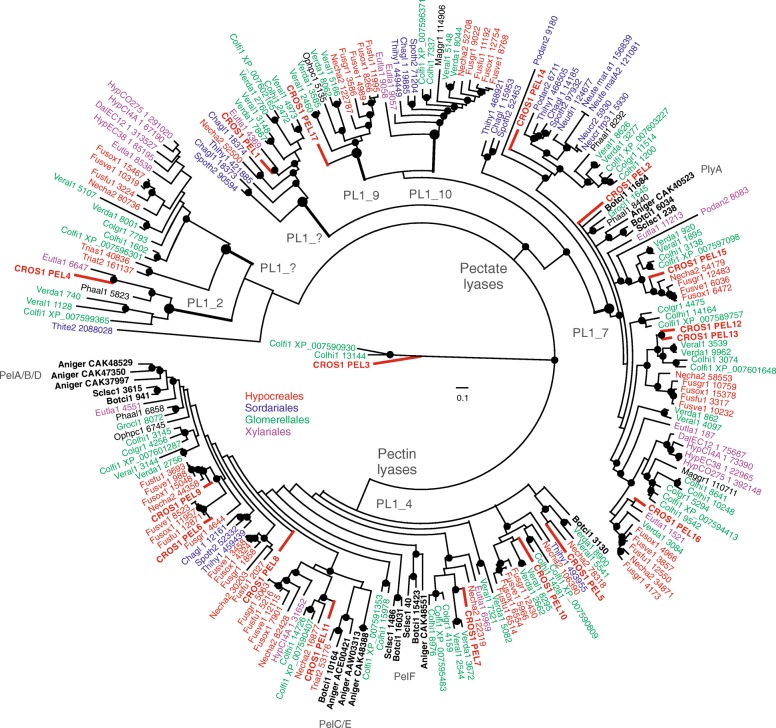
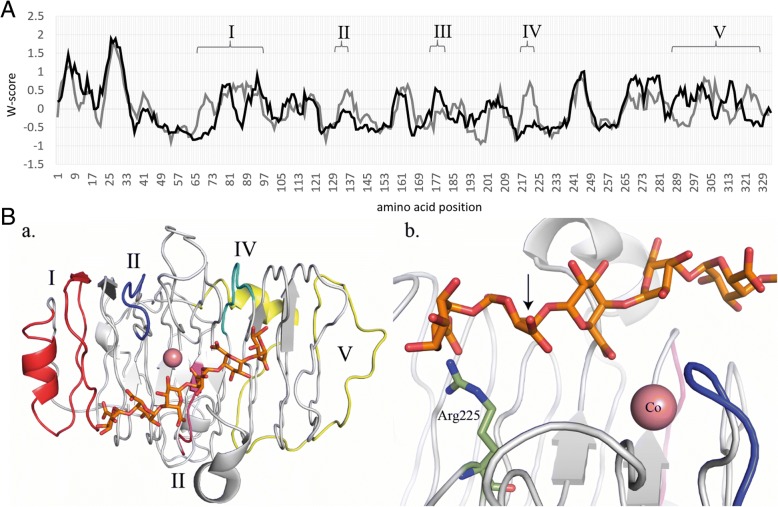
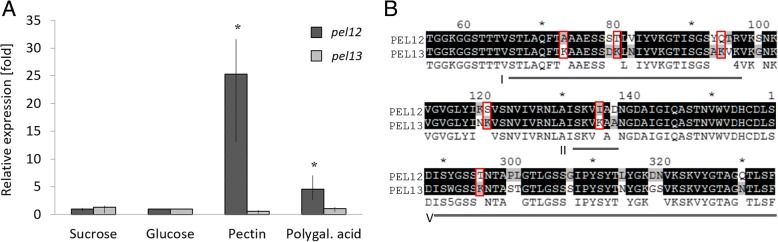
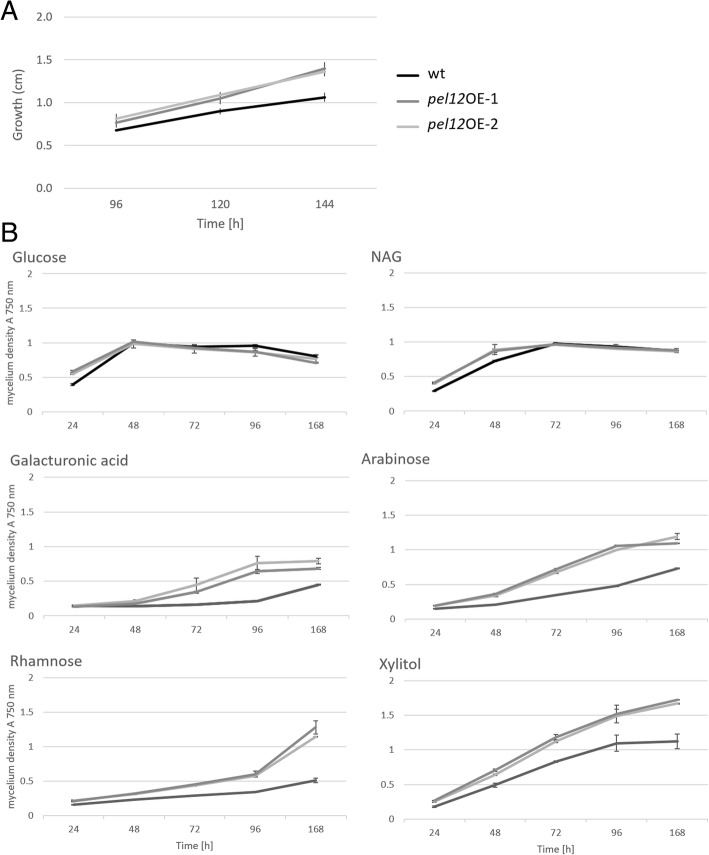
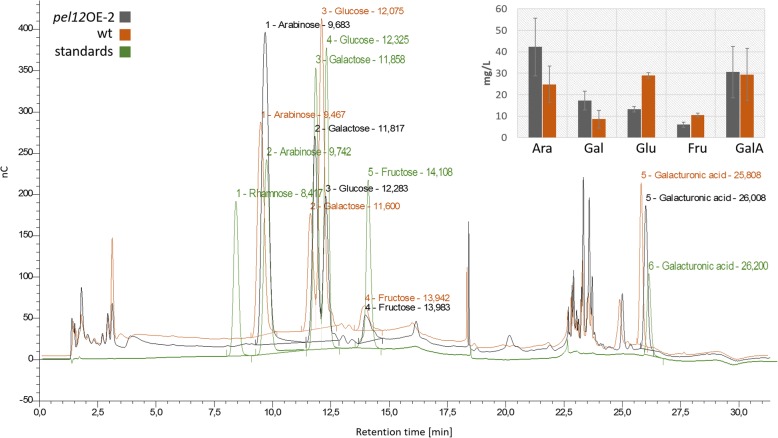
Results: In order to understand the role of the high number of pectin degrading enzymes in Clonostachys, we studied diversity and evolution of the PL1 gene family in C. rosea compared with other Sordariomycetes with varying nutritional life styles. Out of 17 members of C. rosea PL1, we could only detect two to be secreted at acidic pH. One of them, the pectate lyase pel12 gene was found to be strongly induced by pectin and, to a lower degree, by polygalacturonic acid. Heterologous expression of the PEL12 in a PL1-free background of T. reesei revealed direct enzymatic involvement of this protein in utilization of pectin at pH 5 without a requirement for Ca2+. The mutants showed increased utilization of pectin compounds, but did not increase biocontrol ability in detached leaf assay against the plant pathogen Botrytis cinerea compared to the wild type.
Conclusions: In this study, we aimed to gain insight into diversity and evolution of the PL1 gene family in C. rosea and other Sordariomycete species in relation to their nutritional modes. We show that C. rosea PL1 expansion does not correlate with its mycoparasitic nutritional mode and resembles those of strong plant pathogenic fungi. We further investigated regulation, specificity and function of the C. rosea PEL12 and show that this enzyme is directly involved in degradation of pectin and pectin-related compounds, but not in C. rosea biocontrol.
Keywords: Clonostachys rosea; Enzyme; Pectin; Phylogeny; Plant biomass degradation; Trichoderma reesei.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
References
-
- Buchanan BB, Gruissem W, Jones RL, editors. Biochemistry & Molecular Biology of Plants. Rockville: American Society of Plant Physiologists; 2000.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
