

Fibronectin regulates growth factor signaling and cell differentiation in primary lens cells
- PMID: 30404825
- PMCID: PMC6262776
- DOI: 10.1242/jcs.217240
Fibronectin regulates growth factor signaling and cell differentiation in primary lens cells
Abstract
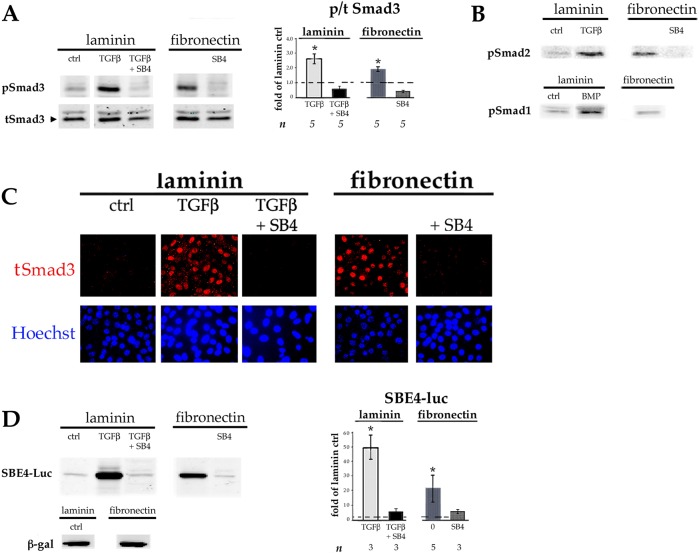
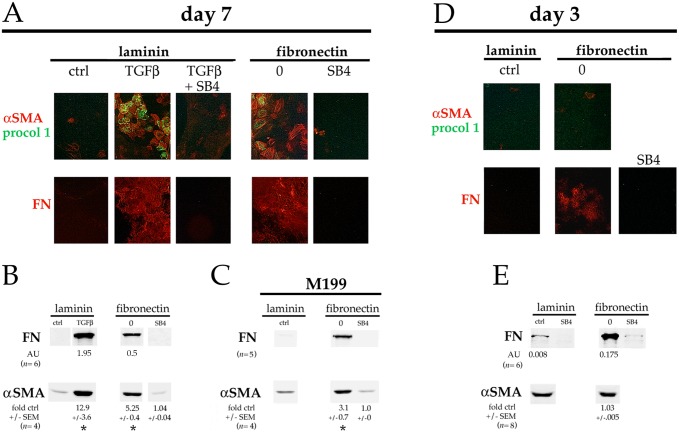
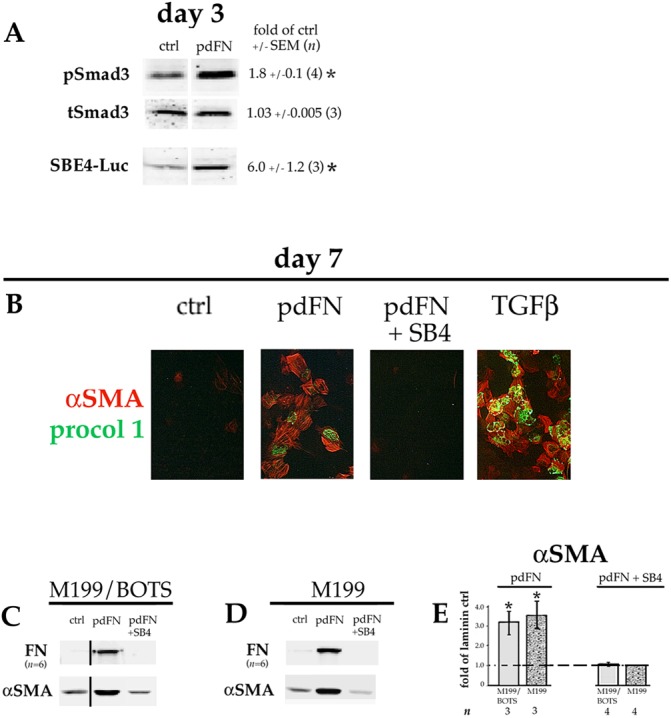
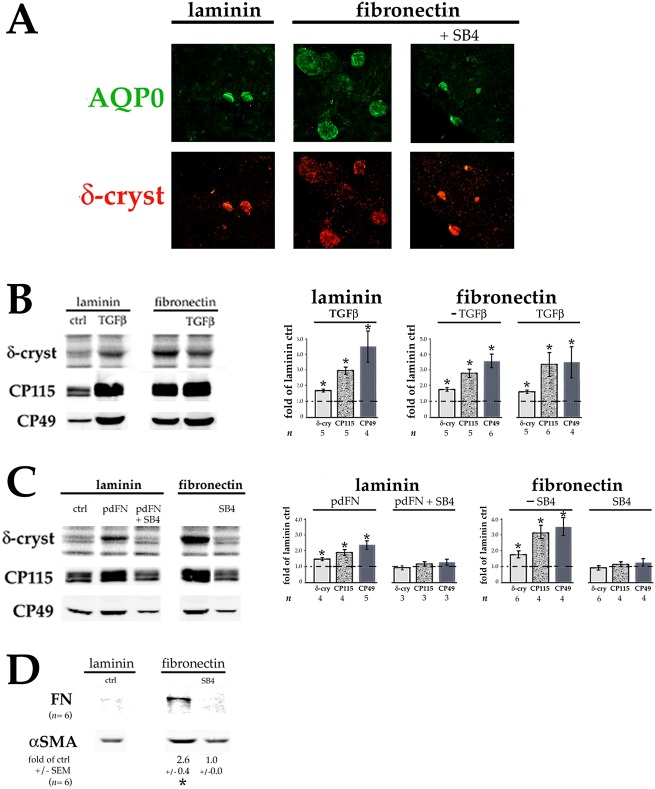
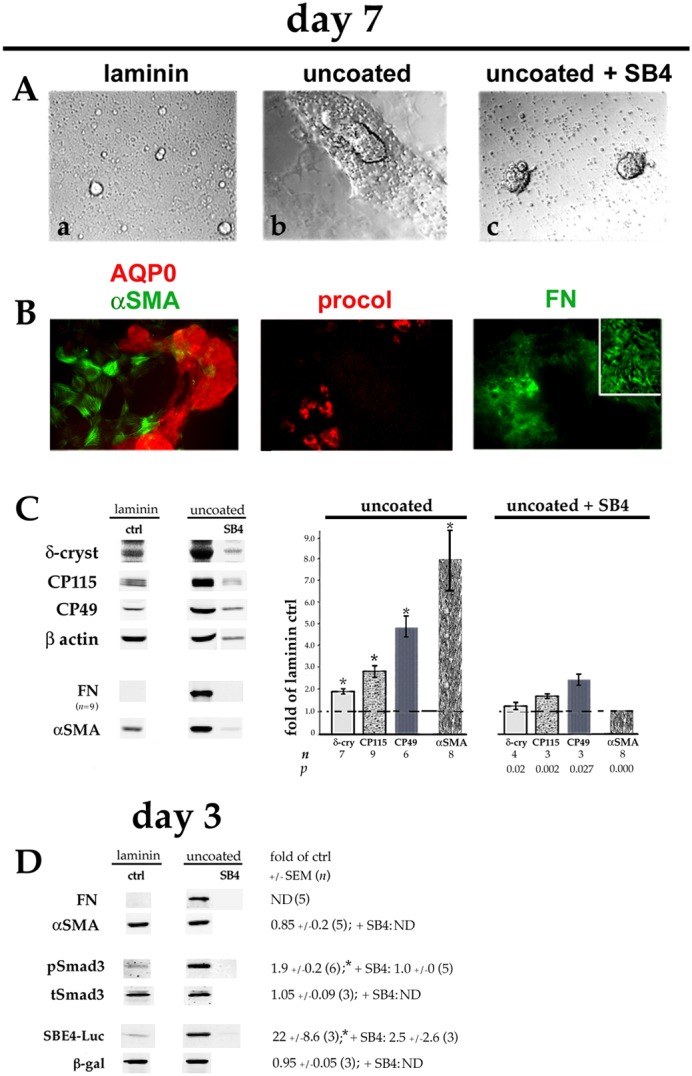
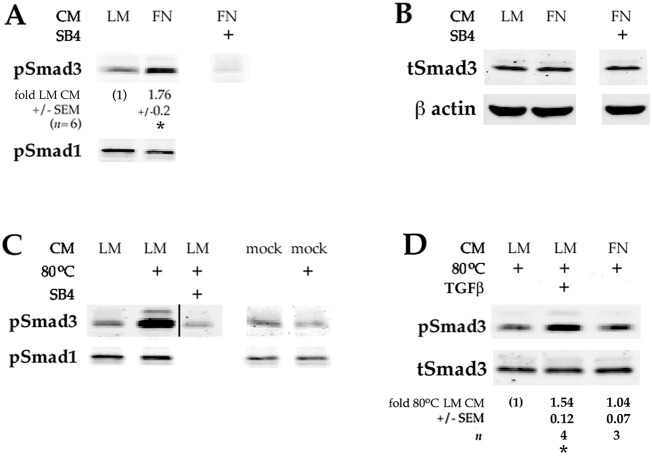
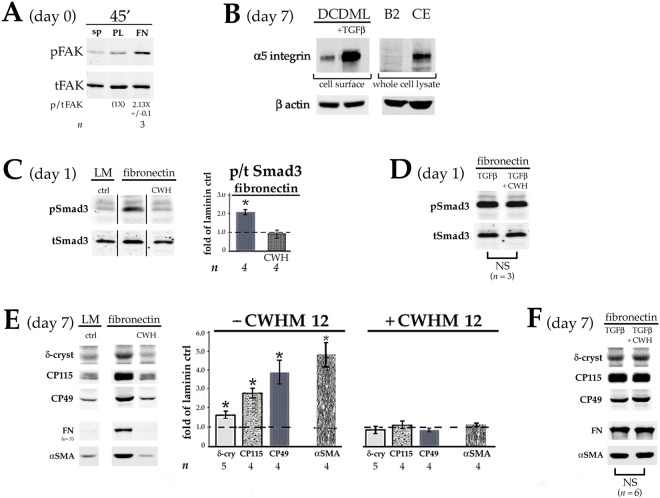
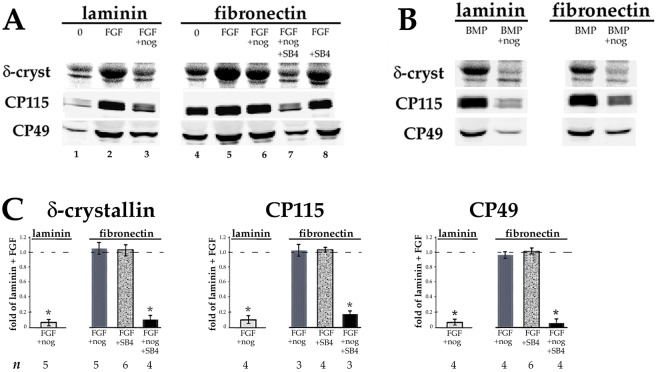
Lens epithelial cells are bound to the lens extracellular matrix capsule, of which laminin is a major component. After cataract surgery, surviving lens epithelial cells are exposed to increased levels of fibronectin, and so we addressed whether fibronectin influences lens cell fate, using DCDML cells as a serum-free primary lens epithelial cell culture system. We found that culturing DCDMLs with plasma-derived fibronectin upregulated canonical TGFβ signaling relative to cells plated on laminin. Fibronectin-exposed cultures also showed increased TGFβ signaling-dependent differentiation into the two cell types responsible for posterior capsule opacification after cataract surgery, namely myofibroblasts and lens fiber cells. Increased TGFβ activity could be identified in the conditioned medium recovered from cells grown on fibronectin. Other experiments showed that plating DCDMLs on fibronectin overcomes the need for BMP in fibroblast growth factor (FGF)-induced lens fiber cell differentiation, a requirement that is restored when endogenous TGFβ signaling is inhibited. These results demonstrate how the TGFβ-fibronectin axis can profoundly affect lens cell fate. This axis represents a novel target for prevention of late-onset posterior capsule opacification, a common but currently intractable complication of cataract surgery.
Keywords: Cataract; Epithelial-mesenchymal transition; Fibrosis; Lens; TGFβ.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
Fibronectin has multifunctional roles in posterior capsular opacification (PCO).Matrix Biol. 2020 Aug;90:79-108. doi: 10.1016/j.matbio.2020.02.004. Epub 2020 Mar 12. Matrix Biol. 2020. PMID: 32173580 Free PMC article.
-
Fibrosis in the lens. Sprouty regulation of TGFβ-signaling prevents lens EMT leading to cataract.Exp Eye Res. 2016 Jan;142:92-101. doi: 10.1016/j.exer.2015.02.004. Epub 2015 May 21. Exp Eye Res. 2016. PMID: 26003864 Free PMC article. Review.
-
Dual function of TGFβ in lens epithelial cell fate: implications for secondary cataract.Mol Biol Cell. 2017 Apr 1;28(7):907-921. doi: 10.1091/mbc.E16-12-0865. Epub 2017 Feb 16. Mol Biol Cell. 2017. PMID: 28209733 Free PMC article.
-
Transforming growth factor-beta-induced epithelial-mesenchymal transition in the lens: a model for cataract formation.Cells Tissues Organs. 2005;179(1-2):43-55. doi: 10.1159/000084508. Cells Tissues Organs. 2005. PMID: 15942192 Review.
-
FGF-2 counteracts loss of TGFbeta affected cells from rat lens explants: implications for PCO (after cataract).Mol Vis. 2004 Jul 22;10:521-32. Mol Vis. 2004. PMID: 15303087
Cited by
-
The Protective Effect of Metformin Use on Early Nd:YAG Laser Capsulotomy.Invest Ophthalmol Vis Sci. 2021 Aug 2;62(10):24. doi: 10.1167/iovs.62.10.24. Invest Ophthalmol Vis Sci. 2021. PMID: 34415985 Free PMC article.
-
TGFβ overcomes FGF-induced transinhibition of EGFR in lens cells to enable fibrotic secondary cataract.Mol Biol Cell. 2024 Jun 1;35(6):ar75. doi: 10.1091/mbc.E24-01-0040. Epub 2024 Apr 10. Mol Biol Cell. 2024. PMID: 38598298 Free PMC article.
-
The Proteomic Landscape of Monocytes in Response to Colorectal Cancer Cells.J Proteome Res. 2024 Sep 6;23(9):4067-4081. doi: 10.1021/acs.jproteome.4c00400. Epub 2024 Aug 6. J Proteome Res. 2024. PMID: 39106312 Free PMC article.
-
Fibronectin has multifunctional roles in posterior capsular opacification (PCO).Matrix Biol. 2020 Aug;90:79-108. doi: 10.1016/j.matbio.2020.02.004. Epub 2020 Mar 12. Matrix Biol. 2020. PMID: 32173580 Free PMC article.
-
ErbBs in Lens Cell Fibrosis and Secondary Cataract.Invest Ophthalmol Vis Sci. 2023 Jul 3;64(10):6. doi: 10.1167/iovs.64.10.6. Invest Ophthalmol Vis Sci. 2023. PMID: 37418274 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
