

Enigma proteins regulate YAP mechanotransduction
- PMID: 30404826
- PMCID: PMC6262774
- DOI: 10.1242/jcs.221788
Enigma proteins regulate YAP mechanotransduction
Abstract
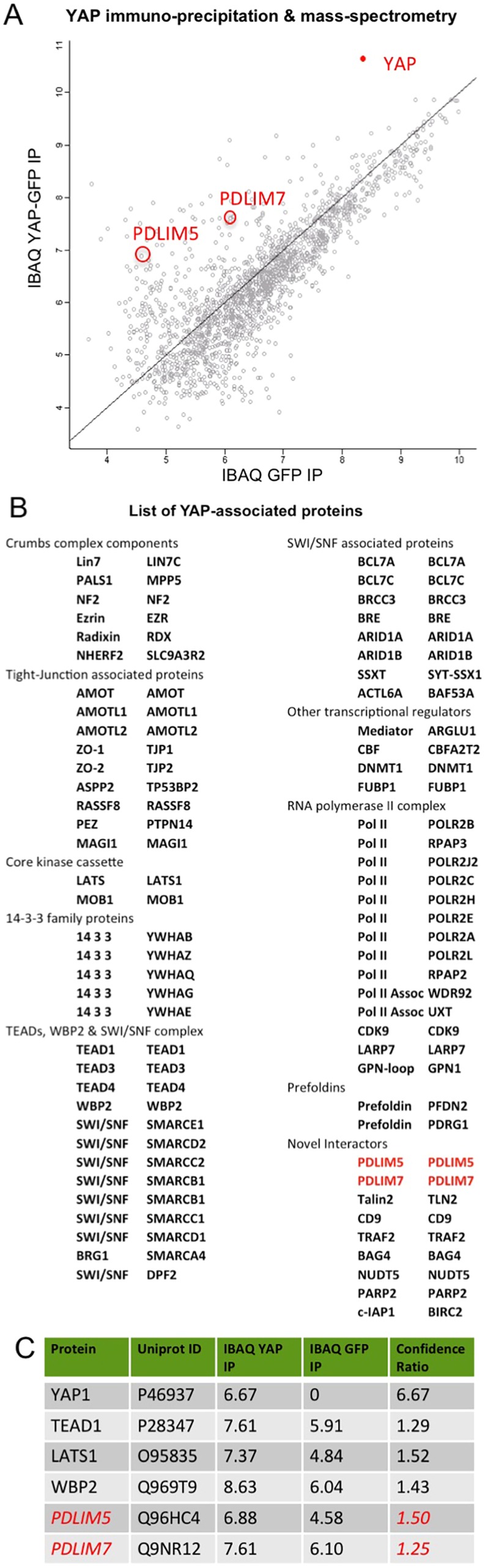
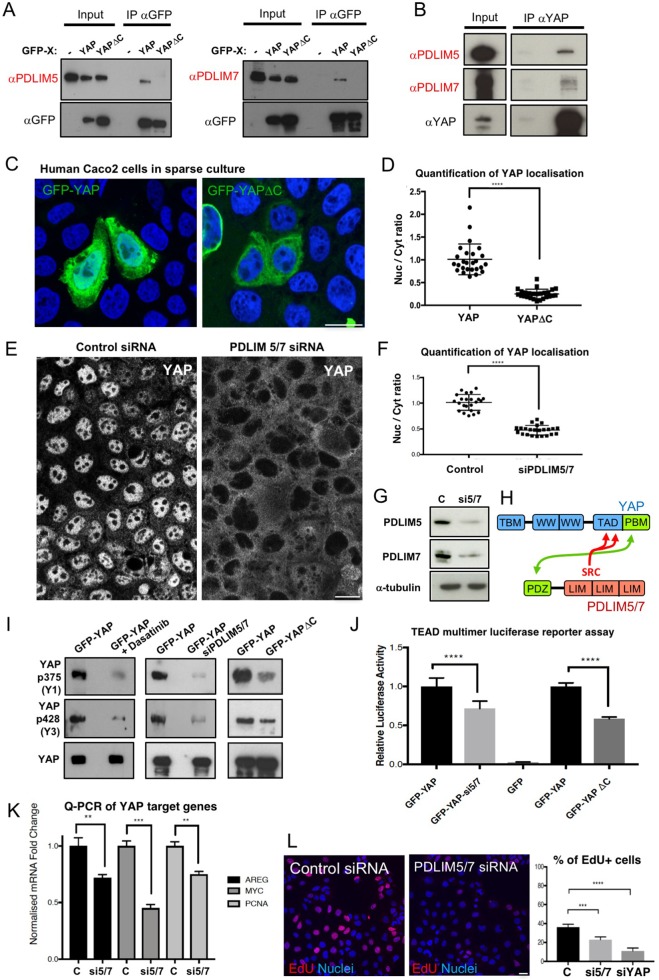
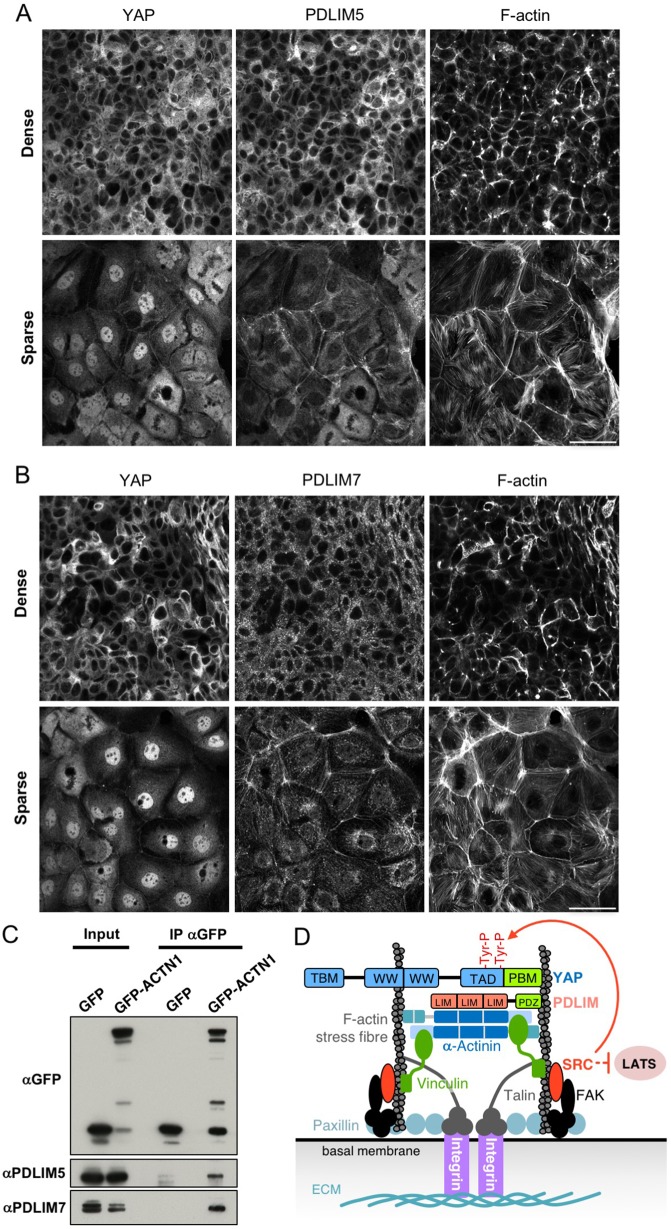
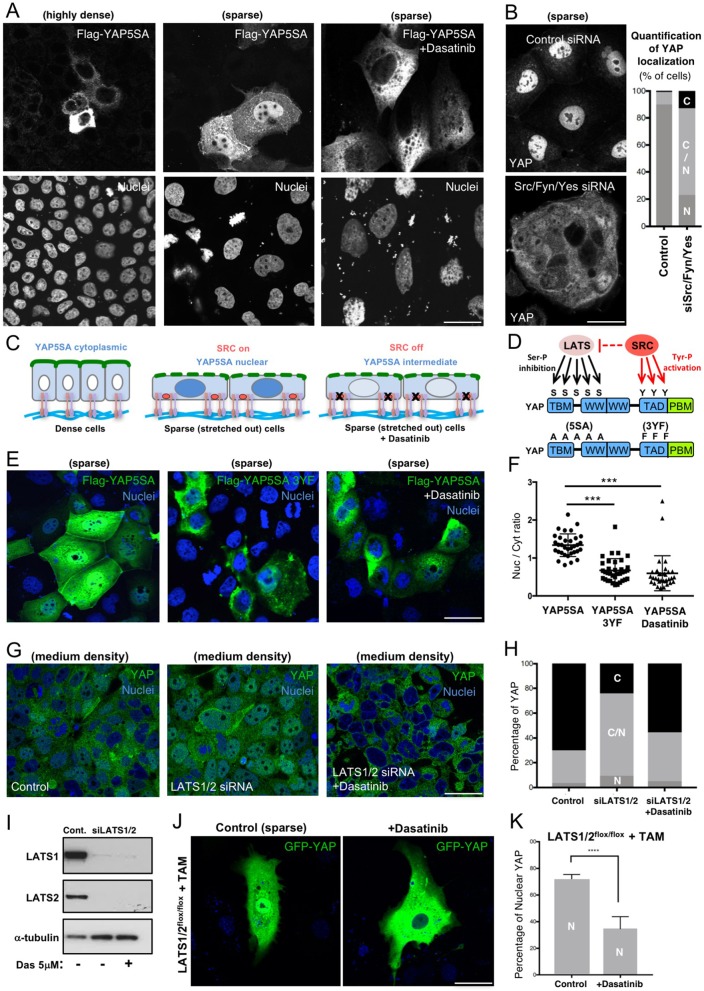
Human cells can sense mechanical stress acting upon integrin adhesions and respond by sending the YAP (also known as YAP1) and TAZ (also known as WWTR1) transcriptional co-activators to the nucleus to drive TEAD-dependent transcription of target genes. How integrin signaling activates YAP remains unclear. Here, we show that integrin-mediated mechanotransduction requires the Enigma and Enigma-like proteins (PDLIM7 and PDLIM5, respectively; denoted for the family of PDZ and LIM domain-containing proteins). YAP binds to PDLIM5 and PDLIM7 (hereafter PDLIM5/7) via its C-terminal PDZ-binding motif (PBM), which is essential for full nuclear localization and activity of YAP. Accordingly, silencing of PDLIM5/7 expression reduces YAP nuclear localization, tyrosine phosphorylation and transcriptional activity. The PDLIM5/7 proteins are recruited from the cytoplasm to integrin adhesions and F-actin stress fibers in response to force by binding directly to the key stress fiber component α-actinin. Thus, forces acting on integrins recruit Enigma family proteins to trigger YAP activation during mechanotransduction.This article has an associated First Person interview with the first author of the paper.
Keywords: Enigma; Hippo; Integrin; Mechanotransduction; PDLIM5; PDLIM7; YAP.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
