

Tumour-targeting bacteria engineered to fight cancer
- PMID: 30405213
- PMCID: PMC6902869
- DOI: 10.1038/s41568-018-0070-z
Tumour-targeting bacteria engineered to fight cancer
Abstract
Recent advances in targeted therapy and immunotherapy have once again raised the hope that a cure might be within reach for many cancer types. Yet, most late-stage cancers are either insensitive to the therapies to begin with or develop resistance later. Therapy with live tumour-targeting bacteria provides a unique option to meet these challenges. Compared with most other therapeutics, the effectiveness of tumour-targeting bacteria is not directly affected by the 'genetic makeup' of a tumour. Bacteria initiate their direct antitumour effects from deep within the tumour, followed by innate and adaptive antitumour immune responses. As microscopic 'robotic factories', bacterial vectors can be reprogrammed following simple genetic rules or sophisticated synthetic bioengineering principles to produce and deliver anticancer agents on the basis of clinical needs. Therapeutic approaches using live tumour-targeting bacteria can either be applied as a monotherapy or complement other anticancer therapies to achieve better clinical outcomes. In this Review, we summarize the potential benefits and challenges of this approach. We discuss how live bacteria selectively induce tumour regression and provide examples to illustrate different ways to engineer bacteria for improved safety and efficacy. Finally, we share our experience and insights on oncology clinical trials with tumour-targeting bacteria, including a discussion of the regulatory issues.
Conflict of interest statement
Figures
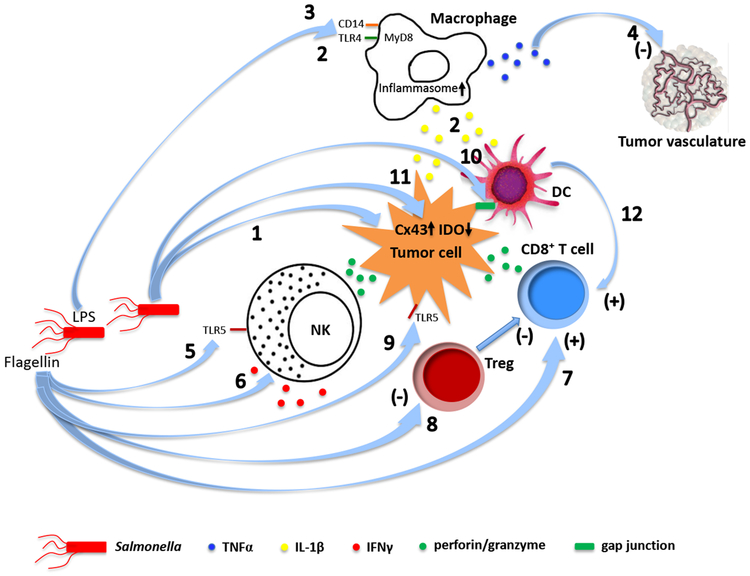
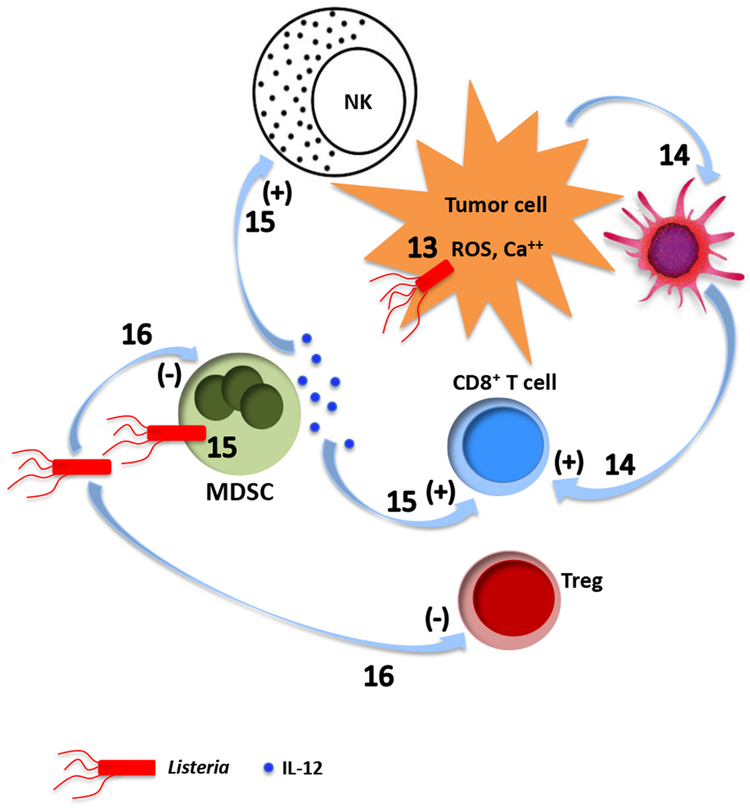
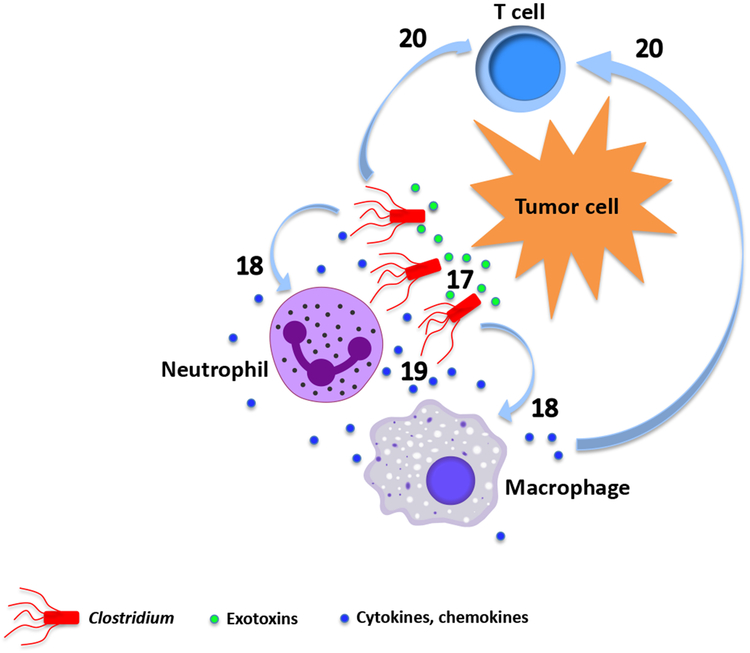
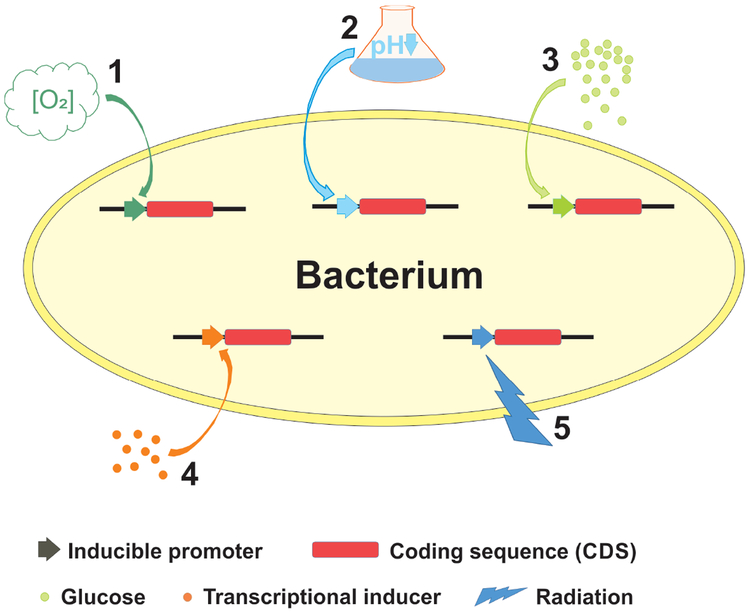
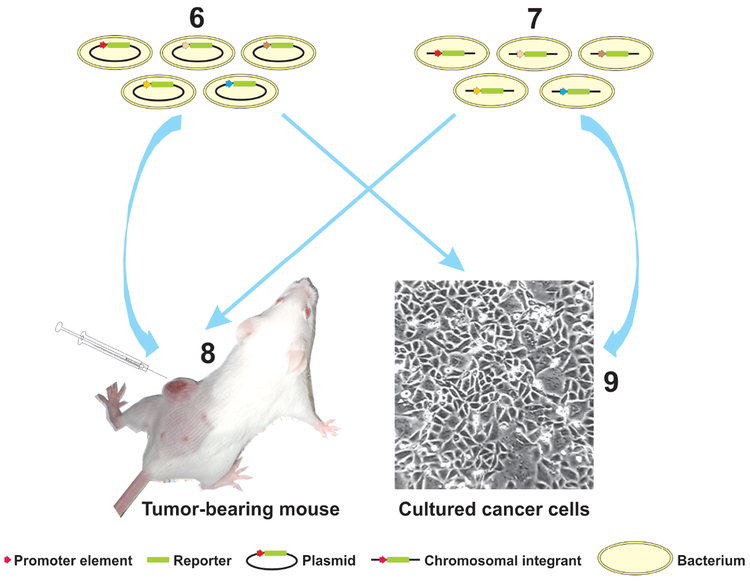
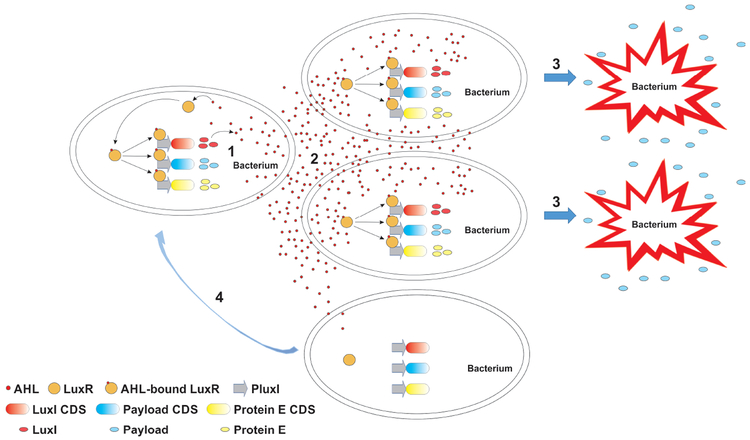
References
-
- Lynch TJ et al. Activating Mutations in the Epidermal Growth Factor Receptor Underlying Responsiveness of Non-Small-Cell Lung Cancer to Gefitinib. N Engl J Med (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
