

Structures of the intermediates of Kok's photosynthetic water oxidation clock
- PMID: 30405241
- PMCID: PMC6485242
- DOI: 10.1038/s41586-018-0681-2
Structures of the intermediates of Kok's photosynthetic water oxidation clock
Abstract
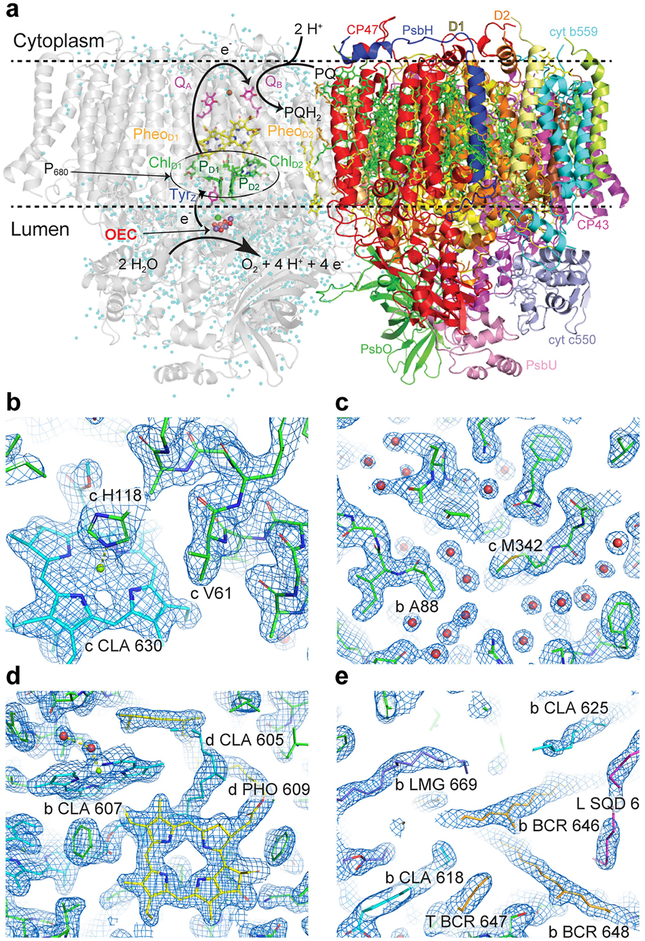
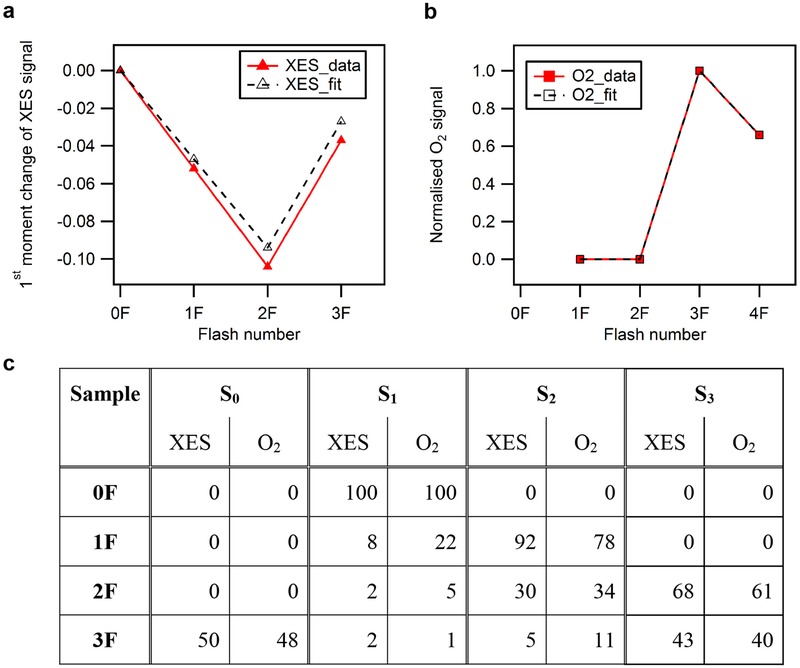
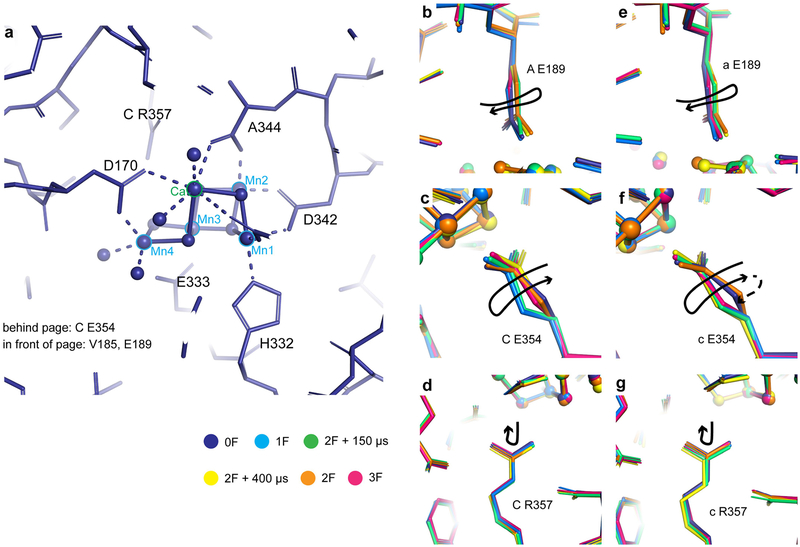
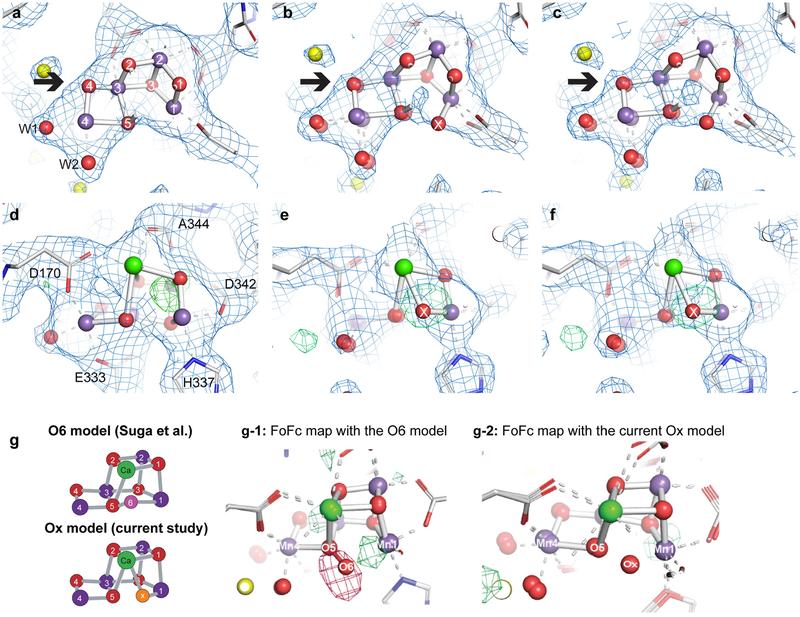
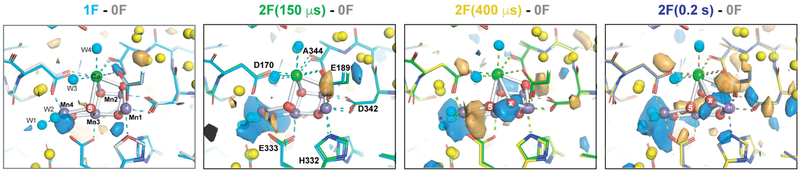
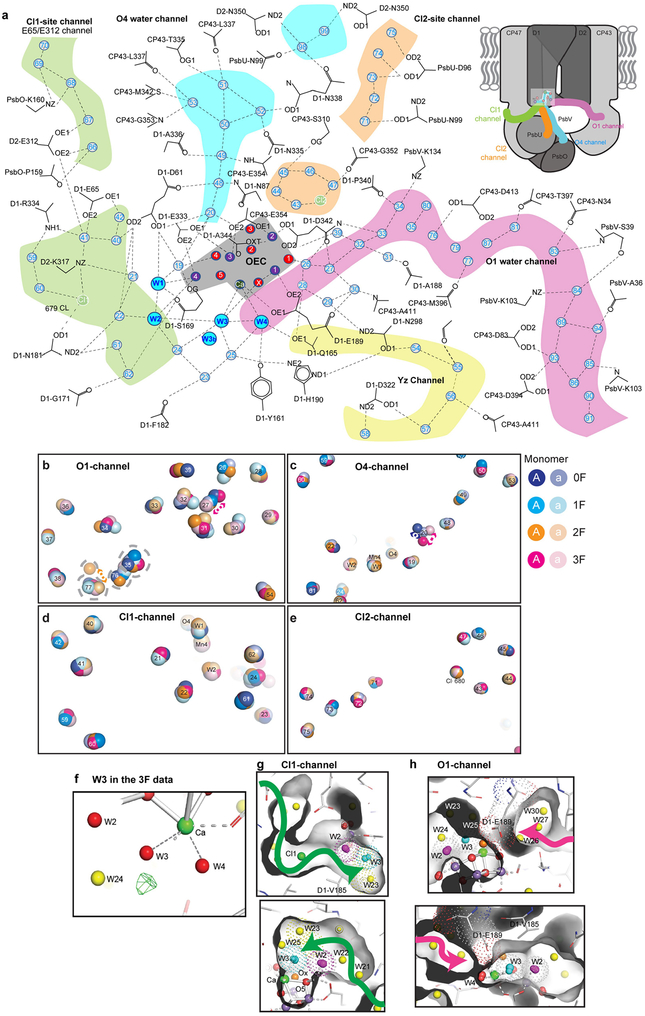
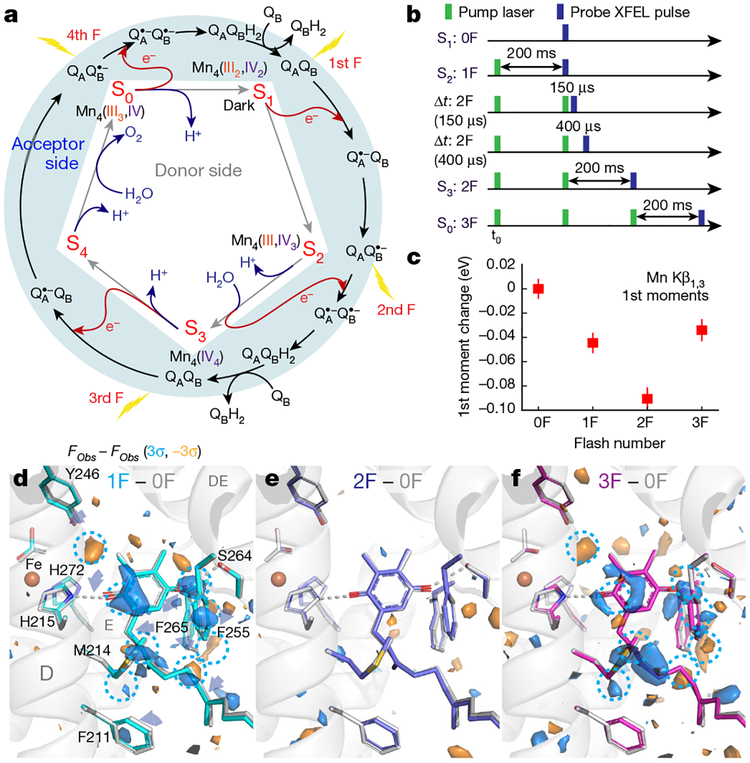
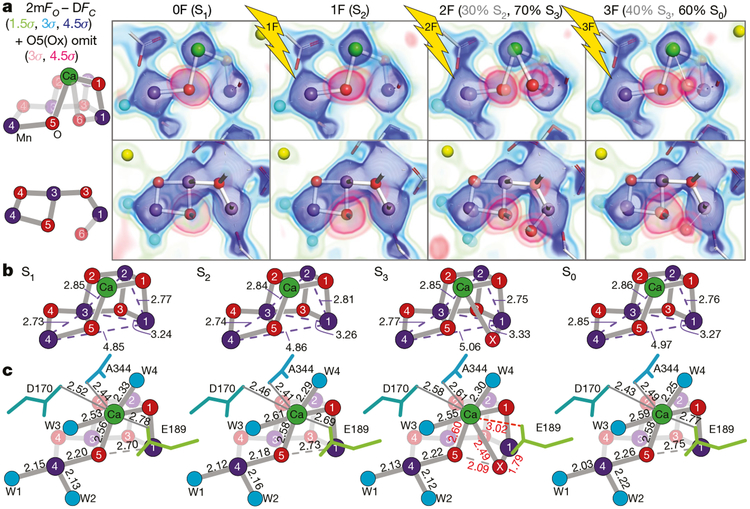
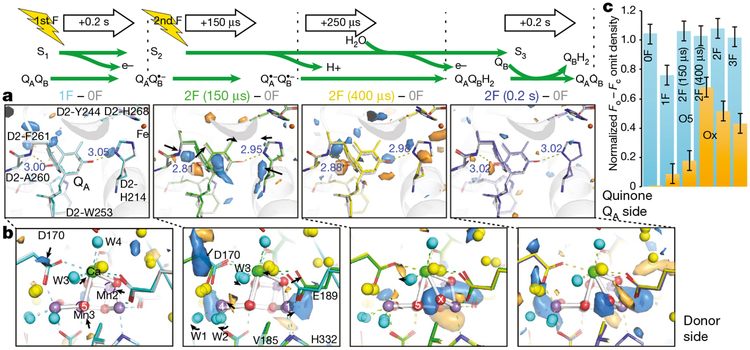
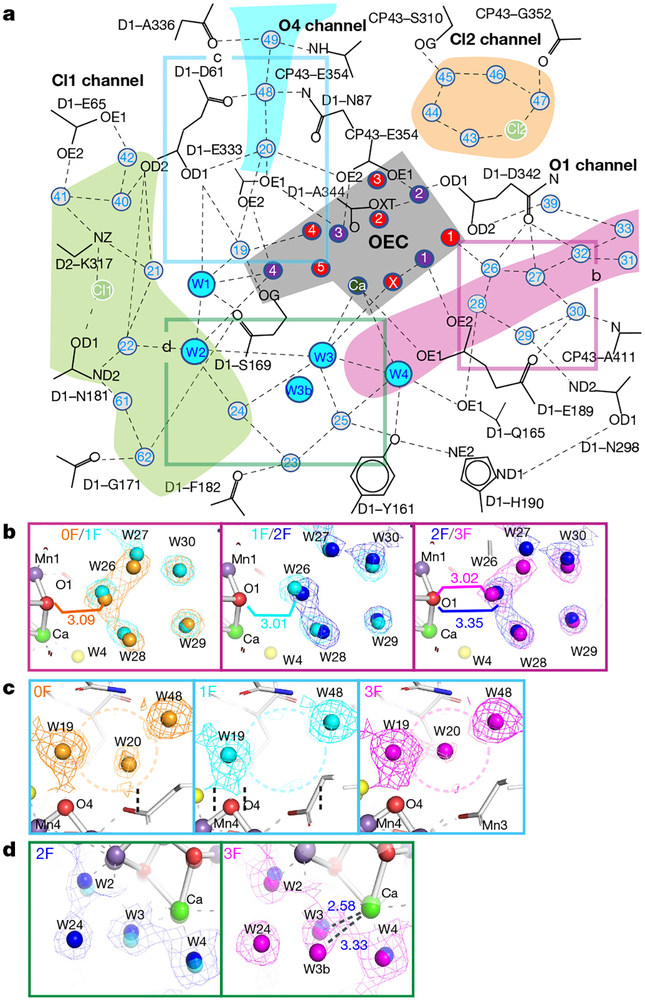
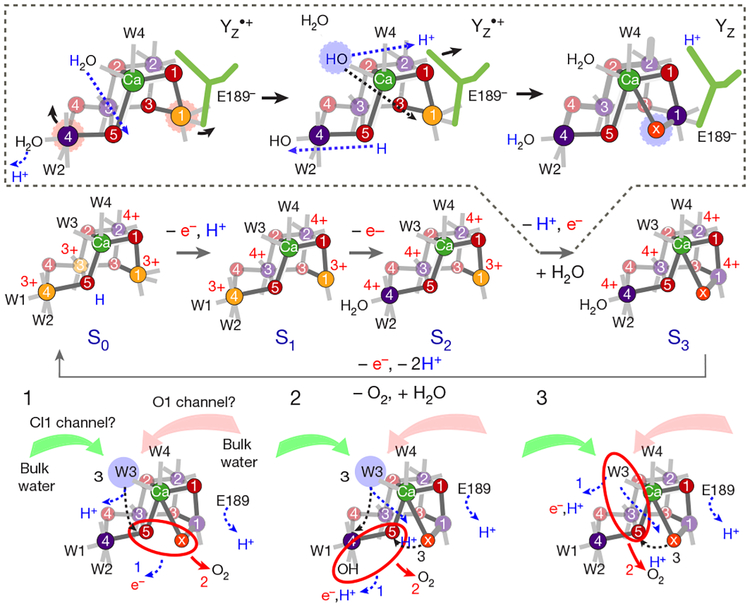
Inspired by the period-four oscillation in flash-induced oxygen evolution of photosystem II discovered by Joliot in 1969, Kok performed additional experiments and proposed a five-state kinetic model for photosynthetic oxygen evolution, known as Kok's S-state clock or cycle1,2. The model comprises four (meta)stable intermediates (S0, S1, S2 and S3) and one transient S4 state, which precedes dioxygen formation occurring in a concerted reaction from two water-derived oxygens bound at an oxo-bridged tetra manganese calcium (Mn4CaO5) cluster in the oxygen-evolving complex3-7. This reaction is coupled to the two-step reduction and protonation of the mobile plastoquinone QB at the acceptor side of PSII. Here, using serial femtosecond X-ray crystallography and simultaneous X-ray emission spectroscopy with multi-flash visible laser excitation at room temperature, we visualize all (meta)stable states of Kok's cycle as high-resolution structures (2.04-2.08 Å). In addition, we report structures of two transient states at 150 and 400 µs, revealing notable structural changes including the binding of one additional 'water', Ox, during the S2→S3 state transition. Our results suggest that one water ligand to calcium (W3) is directly involved in substrate delivery. The binding of the additional oxygen Ox in the S3 state between Ca and Mn1 supports O-O bond formation mechanisms involving O5 as one substrate, where Ox is either the other substrate oxygen or is perfectly positioned to refill the O5 position during O2 release. Thus, our results exclude peroxo-bond formation in the S3 state, and the nucleophilic attack of W3 onto W2 is unlikely.
Conflict of interest statement
Figures

Similar articles
-
Structural evidence for intermediates during O2 formation in photosystem II.Nature. 2023 May;617(7961):629-636. doi: 10.1038/s41586-023-06038-z. Epub 2023 May 3. Nature. 2023. PMID: 37138085 Free PMC article.
-
The O2-Evolving Complex of Photosystem II: Recent Insights from Quantum Mechanics/Molecular Mechanics (QM/MM), Extended X-ray Absorption Fine Structure (EXAFS), and Femtosecond X-ray Crystallography Data.Acc Chem Res. 2017 Jan 17;50(1):41-48. doi: 10.1021/acs.accounts.6b00405. Epub 2016 Dec 21. Acc Chem Res. 2017. PMID: 28001034 Review.
-
Structure of photosystem II and substrate binding at room temperature.Nature. 2016 Dec 15;540(7633):453-457. doi: 10.1038/nature20161. Epub 2016 Nov 21. Nature. 2016. PMID: 27871088 Free PMC article.
-
An oxyl/oxo mechanism for oxygen-oxygen coupling in PSII revealed by an x-ray free-electron laser.Science. 2019 Oct 18;366(6463):334-338. doi: 10.1126/science.aax6998. Epub 2019 Oct 17. Science. 2019. PMID: 31624207
-
An evaluation of structural models for the photosynthetic water-oxidizing complex derived from spectroscopic and X-ray diffraction signatures.J Biol Inorg Chem. 2002 Jan;7(1-2):2-22. doi: 10.1007/s00775-001-0305-3. Epub 2001 Nov 8. J Biol Inorg Chem. 2002. PMID: 11862536 Review.
Cited by
-
Unravelling Mn4Ca cluster vibrations in the S1, S2 and S3 states of the Kok-Joliot cycle of photosystem II.Phys Chem Chem Phys. 2024 Jul 31;26(30):20598-20609. doi: 10.1039/d4cp01307g. Phys Chem Chem Phys. 2024. PMID: 39037338 Free PMC article.
-
Capturing the sequence of events during the water oxidation reaction in photosynthesis using XFELs.FEBS Lett. 2023 Jan;597(1):30-37. doi: 10.1002/1873-3468.14527. Epub 2022 Nov 15. FEBS Lett. 2023. PMID: 36310373 Free PMC article. Review.
-
Interplay of two low-barrier hydrogen bonds in long-distance proton-coupled electron transfer for water oxidation.PNAS Nexus. 2023 Dec 7;2(12):pgad423. doi: 10.1093/pnasnexus/pgad423. eCollection 2023 Dec. PNAS Nexus. 2023. PMID: 38130665 Free PMC article.
-
Characterization of the Rate-Limiting Steps in the Dark-To-Light Transitions of Closed Photosystem II: Temperature Dependence and Invariance of Waiting Times during Multiple Light Reactions.Int J Mol Sci. 2022 Dec 21;24(1):94. doi: 10.3390/ijms24010094. Int J Mol Sci. 2022. PMID: 36613535 Free PMC article.
-
In situ observation of picosecond polaron self-localisation in α-Fe2O3 photoelectrochemical cells.Nat Commun. 2019 Sep 3;10(1):3962. doi: 10.1038/s41467-019-11767-9. Nat Commun. 2019. PMID: 31481691 Free PMC article.
References
-
- Kok B, Forbush B & McGloin M Cooperation of charges in photosynthetic O2 evolution—I. A linear four step mechanism. Photochem. Photobiol 11, 457–475 (1970). - PubMed
-
- Joliot P, Barbieri G & Chabaud R A new model of photochemical centers in system-2. Photochem. Photobiol 10, 309–329 (1969).
-
- Hillier W & Wydrzynski T 18O-water exchange in photosystem II: substrate binding and intermediates of the water splitting cycle. Coord. Chem. Rev 252, 306–317 (2008).
-
- Cox N & Messinger J Reflections on substrate water and dioxygen formation. Biochim. Biophys. Acta 1827, 1020–1030 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- GM117126/GM/NIGMS NIH HHS/United States
- P30 GM124169/GM/NIGMS NIH HHS/United States
- GM116423-02/GM/NIGMS NIH HHS/United States
- R01 GM126289/GM/NIGMS NIH HHS/United States
- R56 GM055302/GM/NIGMS NIH HHS/United States
- R01 GM110501/GM/NIGMS NIH HHS/United States
- F32 GM116423/GM/NIGMS NIH HHS/United States
- P01 GM063210/GM/NIGMS NIH HHS/United States
- R01 GM124149/GM/NIGMS NIH HHS/United States
- P41 GM103393/GM/NIGMS NIH HHS/United States
- R01 GM055302/GM/NIGMS NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 GM117126/GM/NIGMS NIH HHS/United States
