

Influence of Magnitude and Duration of Altered Gravity and Readaptation to 1 g on the Structure and Function of the Utricle in Toadfish, Opsanus tau
- PMID: 30405430
- PMCID: PMC6204554
- DOI: 10.3389/fphys.2018.01469
Influence of Magnitude and Duration of Altered Gravity and Readaptation to 1 g on the Structure and Function of the Utricle in Toadfish, Opsanus tau
Abstract
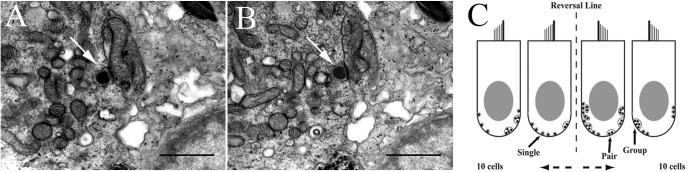
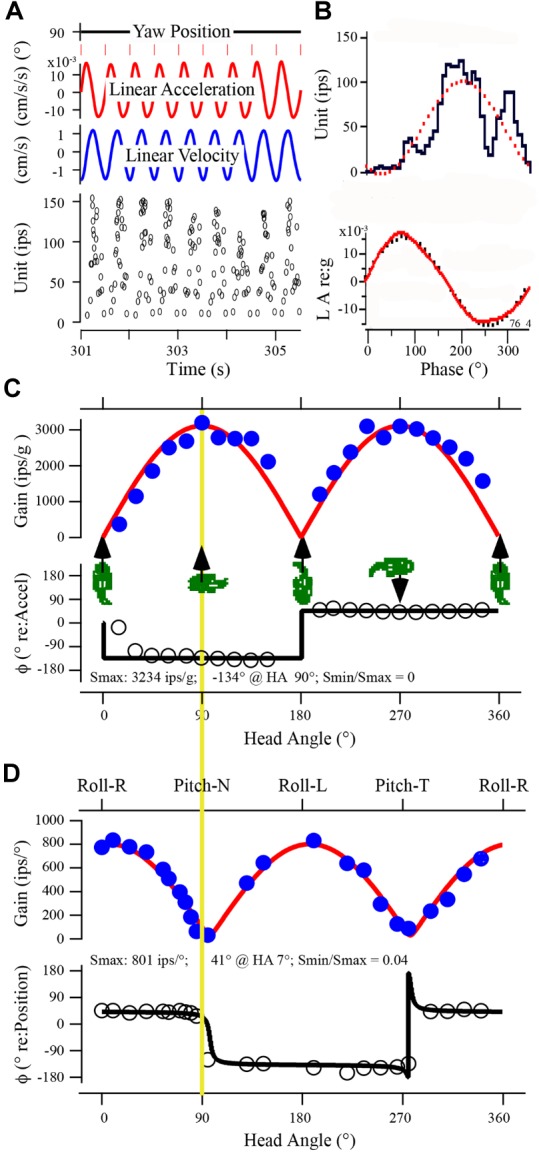
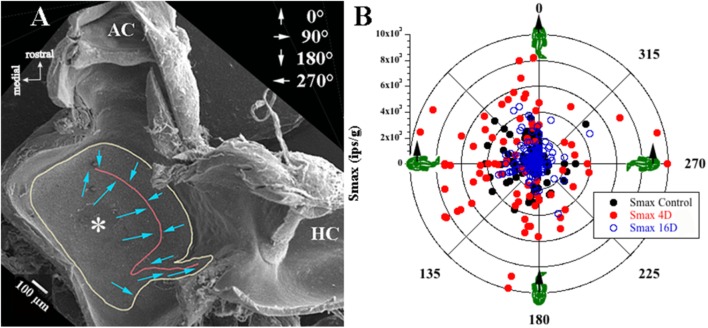
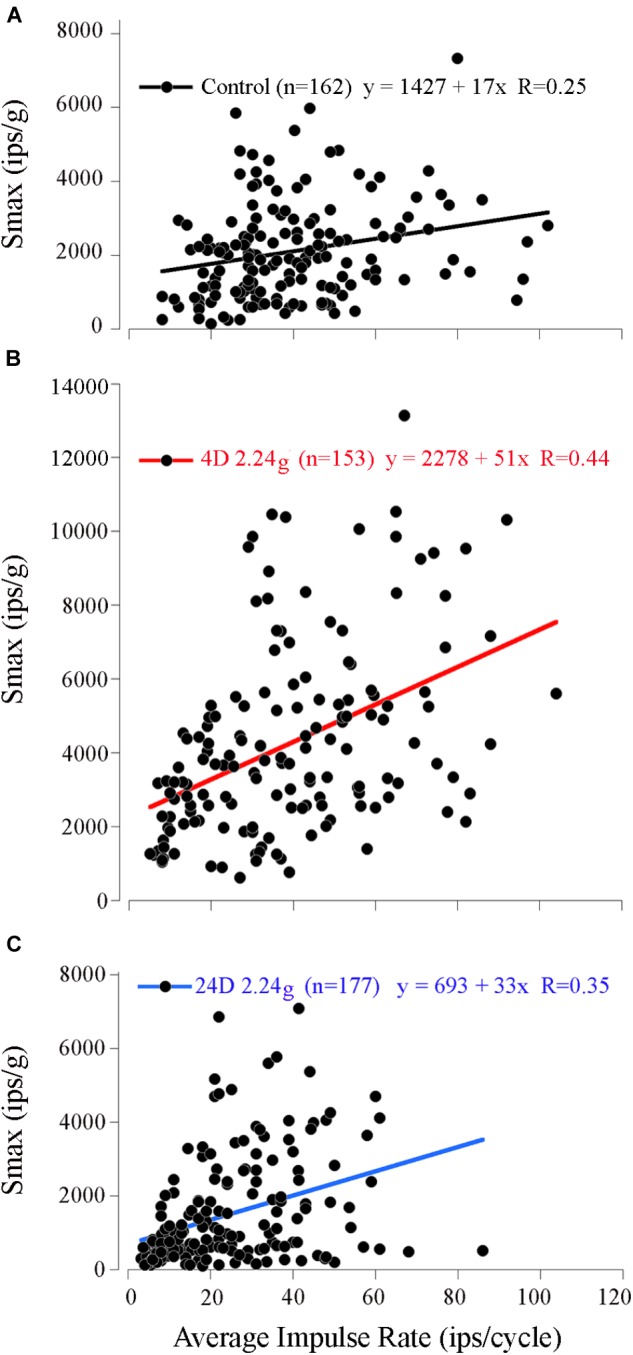
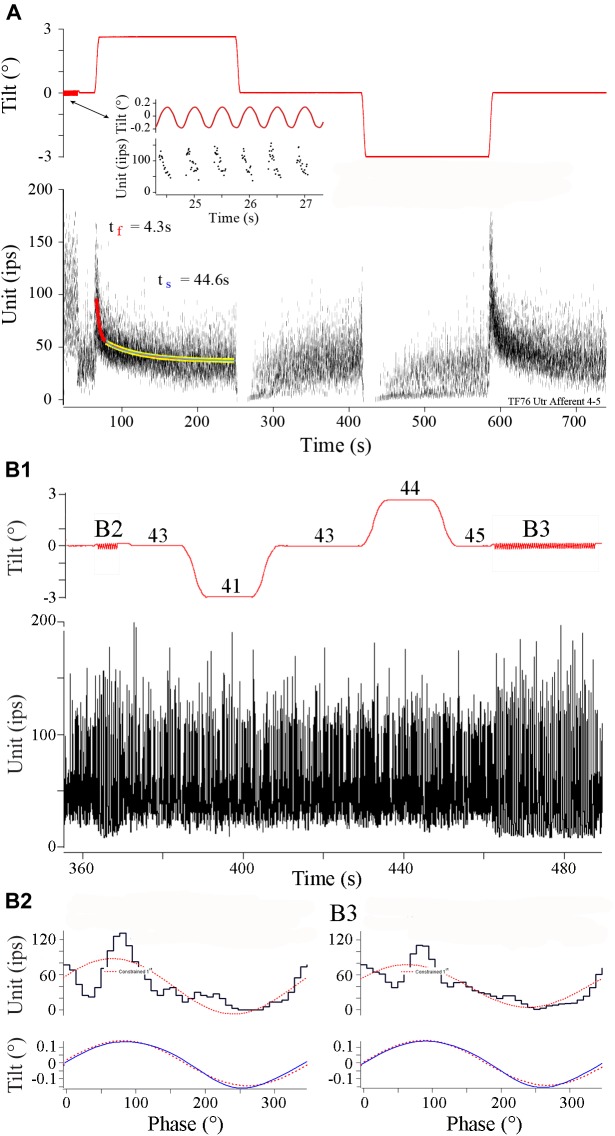
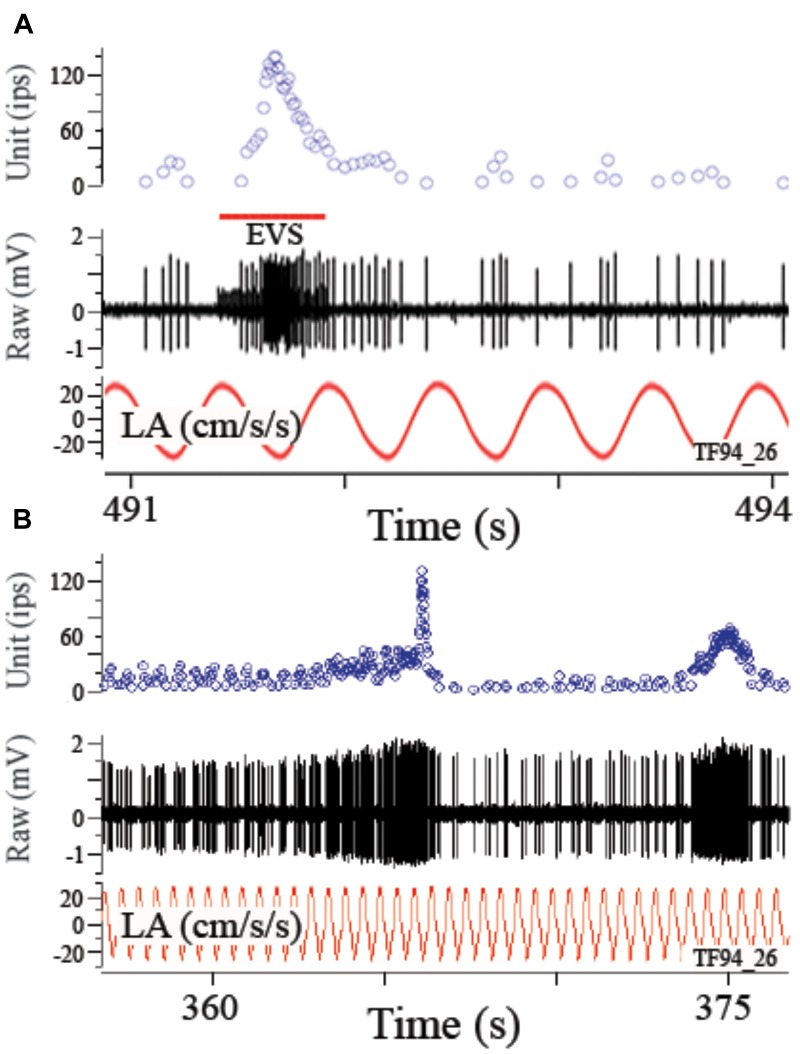
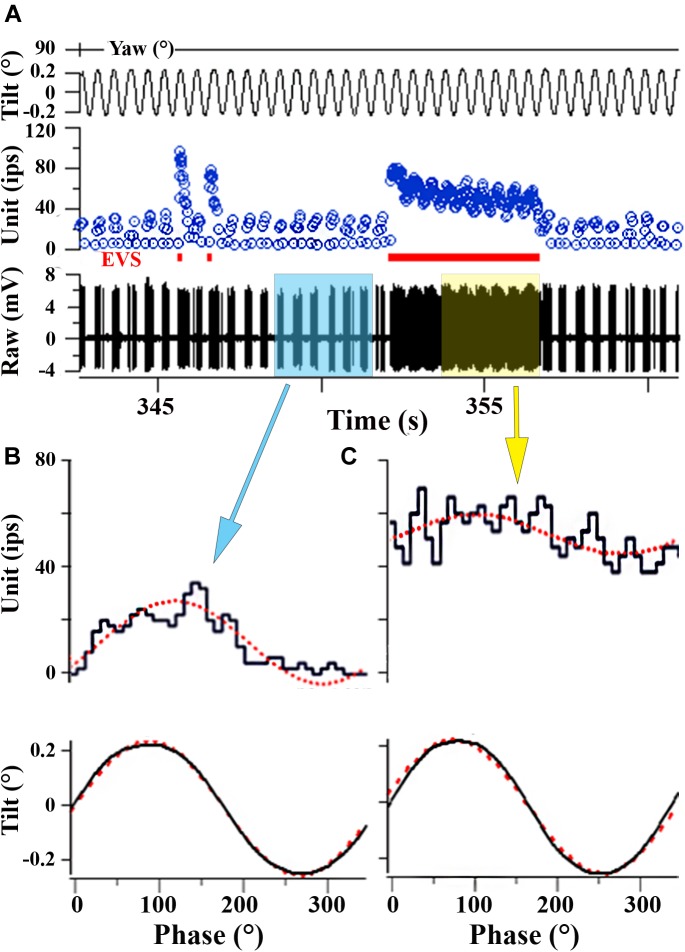
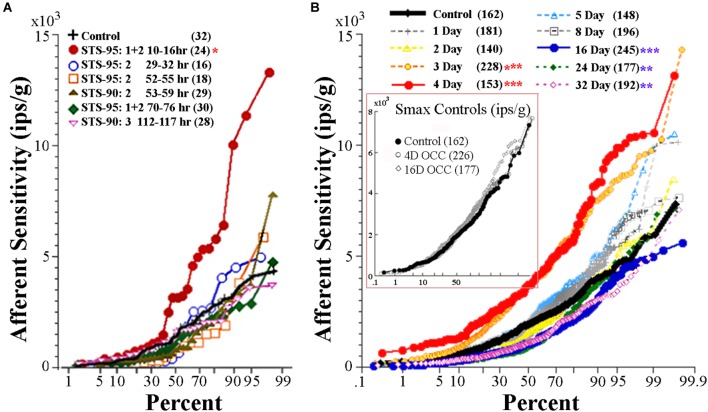
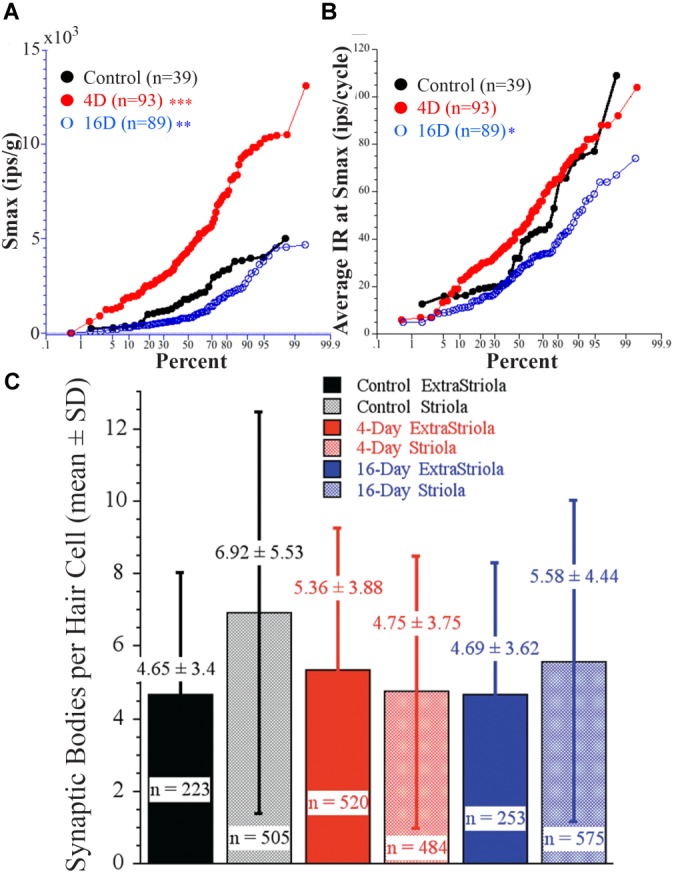
Gravity has remained constant during animal evolution and the neural sensory systems detecting acceleration forces have remained remarkably conserved among vertebrates. The utricular organ senses the sum of inertial force due to head translation and head tilt relative to gravitational vertical. Change in gravitational force would be expected to have profound effects on how an organism maintains equilibrium. We characterize the physiology of utricular afferents to applied accelerations in the oyster toadfish, Opsanus tau, in normal 1 g to establish benchmarks, after 1-32-day exposures to 2.24 g (resultant) via centrifugation (hypergravity, HG), after 4- and 16-day exposures to 1.12 g (resultant), and following 1-8 days recovery to HG exposures to study re-adaptation to 1 g. Afferents were also examined during activation of efferent vestibular pathway. Centrifugation at 2.24 g included 228°/s constant angular velocity component, and thus horizontal canal afferent responses to yaw rotation were recorded as an internal control in each fish. Afferents studied after 228°/s rotation for 4 and 16 days without centripetal acceleration, called On-Center-Control, were indistinguishable from their control counterparts. Principal response to HG was an adjustment of afferent sensitivity as a function of magnitude and duration of exposure: an initial robust increase at 3-4 days followed by a significant decrease from 16 to 32 days. Initial increase observed after 4 days of HG took >4 days in 1 g to recover, and the decrease observed after 16 days of HG took >2 days to readapt to 1 g. Hair cells in striola and medial extrastriola macula regions were serially reconstructed in 3D from thin sections using transmission electron microscopy in control fish and fish exposed to 4 and 16 days of HG. Despite the highly significant differences in afferent physiology, synaptic body counts quantified in the same fish were equivalent in their inter-animal variability and averages. No clear role of the efferent pathway as a feedback mechanism regulating afferent behavior to HG was found. Transfer from 1 g to HG imparts profound effects on gravitational sensitivity of utricular afferents and the accompanying transfer from the HG back to the 1 g resembles in part (as an analog) the transfer from 1 g to the micrograms.
Keywords: acceleration; afferents; centrifugation; electrophysiology; gravity; hair cells; serial electron microscopy reconstruction; spaceflight.
Figures


Similar articles
-
Otolith adaptive responses to altered gravity.Neurosci Biobehav Rev. 2021 Mar;122:218-228. doi: 10.1016/j.neubiorev.2020.10.025. Epub 2020 Nov 3. Neurosci Biobehav Rev. 2021. PMID: 33152424 Review.
-
Morphology of the utricular otolith organ in the toadfish, Opsanus tau.J Comp Neurol. 2018 Jun 15;526(9):1571-1588. doi: 10.1002/cne.24429. Epub 2018 Mar 23. J Comp Neurol. 2018. PMID: 29524209 Free PMC article.
-
Neural readaptation to Earth's gravity following return from space.J Neurophysiol. 2001 Oct;86(4):2118-22. doi: 10.1152/jn.2001.86.4.2118. J Neurophysiol. 2001. PMID: 11600668
-
Response of toadfish (Opsanus tau) utricular afferents to multimodal inputs.J Neurophysiol. 2022 Aug 1;128(2):364-377. doi: 10.1152/jn.00483.2021. Epub 2022 Jul 13. J Neurophysiol. 2022. PMID: 35830608
-
Gravity or translation: central processing of vestibular signals to detect motion or tilt.J Vestib Res. 2003;13(4-6):245-53. J Vestib Res. 2003. PMID: 15096668 Review.
Cited by
-
Reproductive state modulates utricular auditory sensitivity in a vocal fish.J Neurophysiol. 2022 Nov 1;128(5):1344-1354. doi: 10.1152/jn.00315.2022. Epub 2022 Oct 26. J Neurophysiol. 2022. PMID: 36286323 Free PMC article.
-
Peripheral vestibular plasticity vs central compensation: evidence and questions.J Neurol. 2019 Sep;266(Suppl 1):27-32. doi: 10.1007/s00415-019-09388-9. Epub 2019 May 27. J Neurol. 2019. PMID: 31134376 Review.
References
LinkOut - more resources
Full Text Sources