

A Metalloprotease Homolog Venom Protein From a Parasitoid Wasp Suppresses the Toll Pathway in Host Hemocytes
- PMID: 30405599
- PMCID: PMC6206080
- DOI: 10.3389/fimmu.2018.02301
A Metalloprotease Homolog Venom Protein From a Parasitoid Wasp Suppresses the Toll Pathway in Host Hemocytes
Abstract
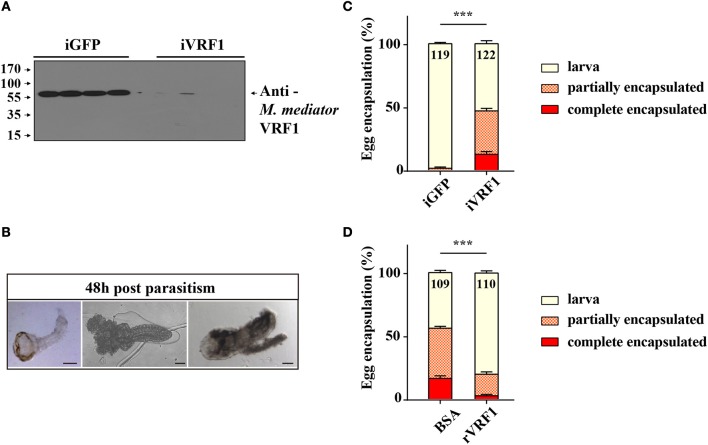
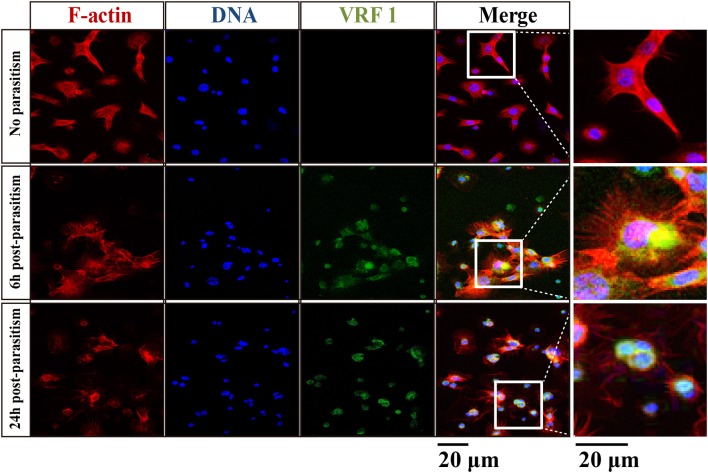
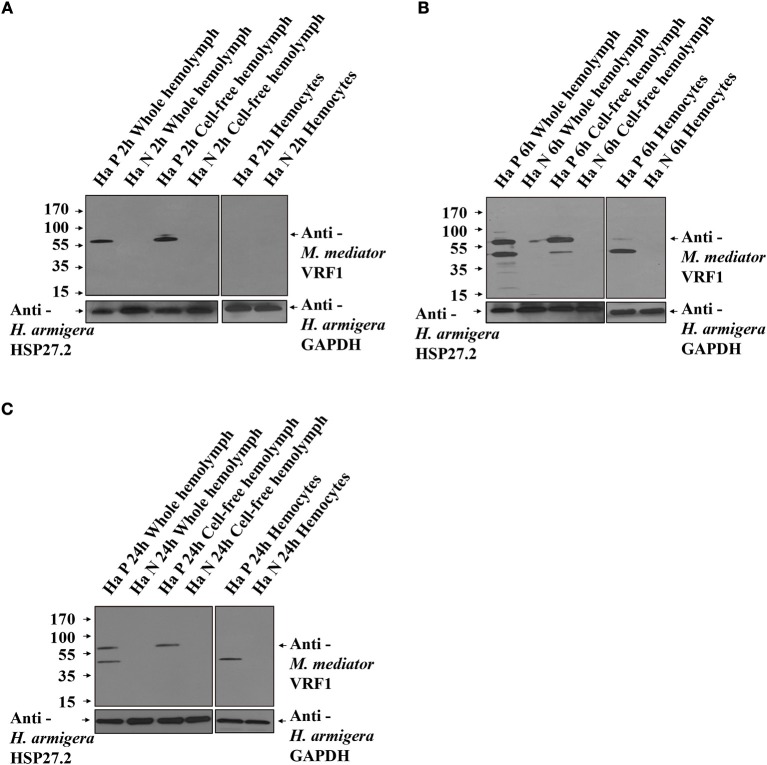
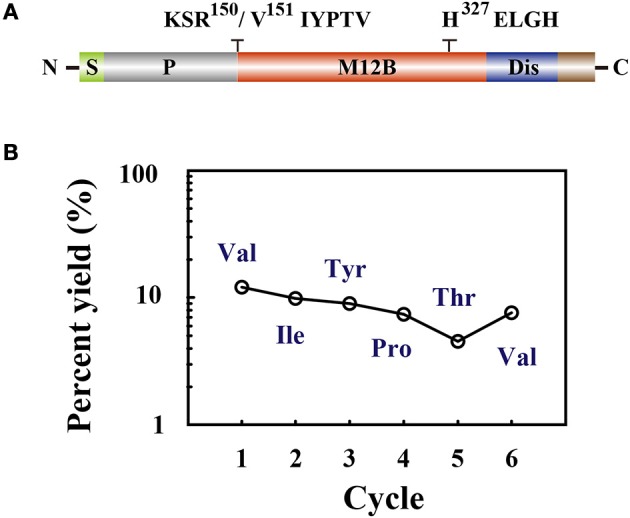
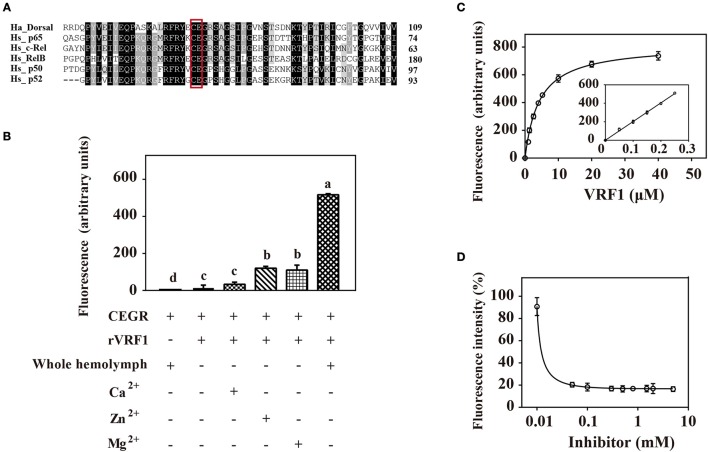
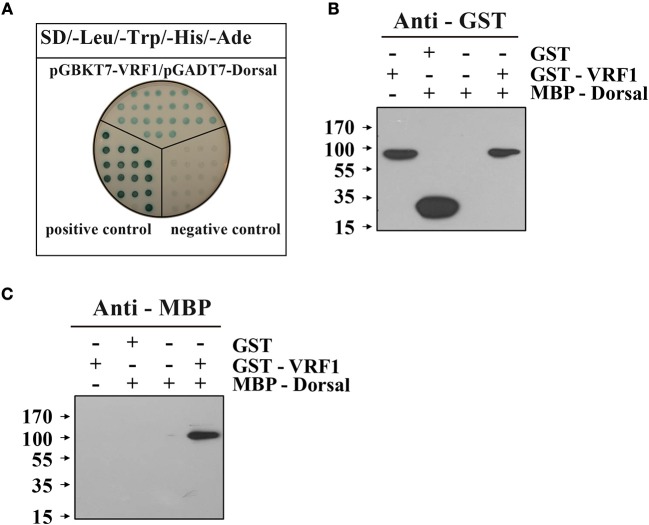
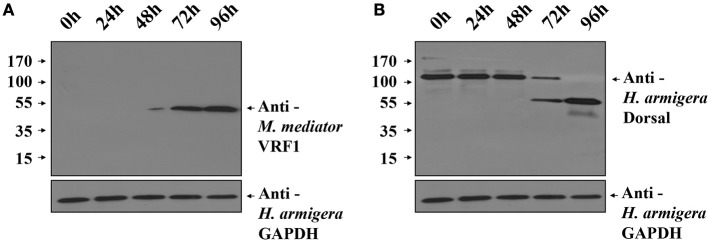
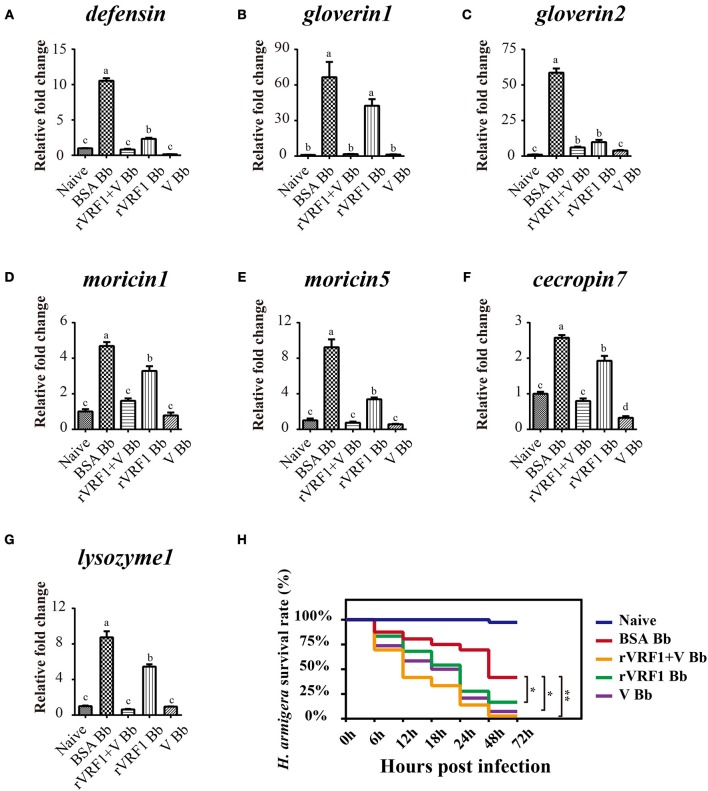
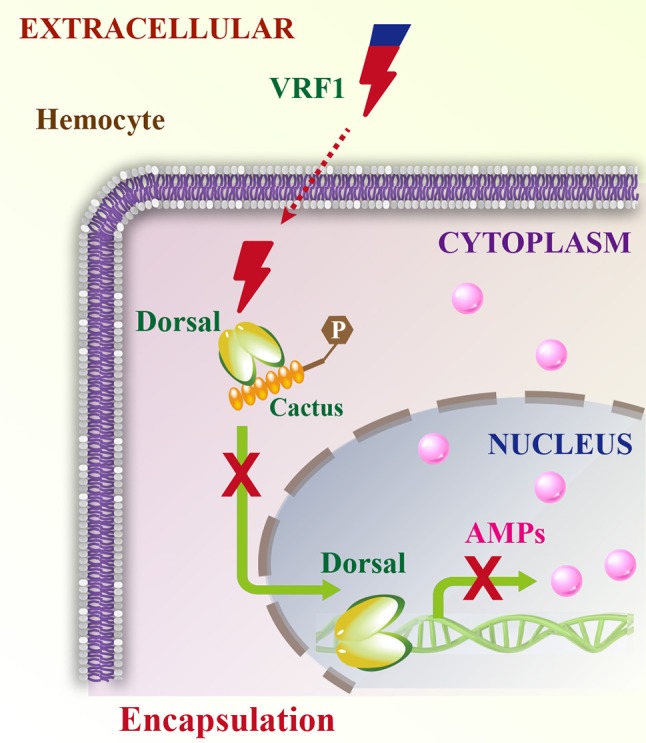
Parasitoid wasps depend on a variety of maternal virulence factors to ensure successful parasitism. Encapsulation response carried out by host hemocytes is one of the major host immune responses toward limiting endoparasitoid wasp offspring production. We found that VRF1, a metalloprotease homolog venom protein identified from the endoparasitoid wasp, Microplitis mediator, could modulate egg encapsulation in its host, the cotton bollworm, Helicoverpa armigera. Here, we show that the VRF1 proenzyme is cleaved after parasitism, and that the C-terminal fragment containing the catalytic domain enters host hemocytes 6 h post-parasitism. Furthermore, using yeast two-hybrid and pull-down assays, VRF1 is shown to interact with the H. armigera NF-κB factor, Dorsal. We also show that overexpressed of VRF1 in an H. armigera cell line cleaved Dorsal in vivo. Taken together, our results have revealed a novel mechanism by which a component of endoparasitoid wasp venom interferes with the Toll signaling pathway in the host hemocytes.
Keywords: Helicoverpa armigera; Microplitis mediator; metalloprotease; toll pathway; venom.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources