

Microglia Enhance Synapse Activity to Promote Local Network Synchronization
- PMID: 30406198
- PMCID: PMC6220592
- DOI: 10.1523/ENEURO.0088-18.2018
Microglia Enhance Synapse Activity to Promote Local Network Synchronization
Abstract
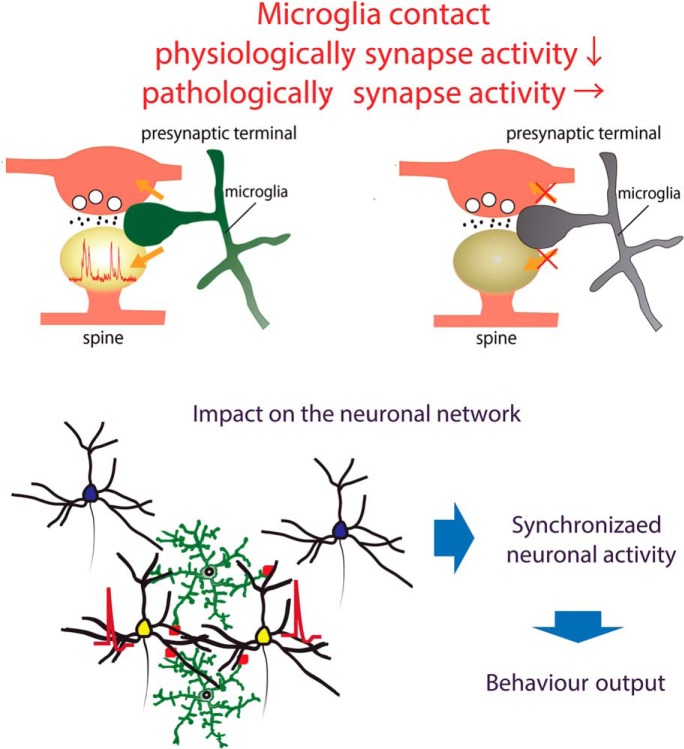
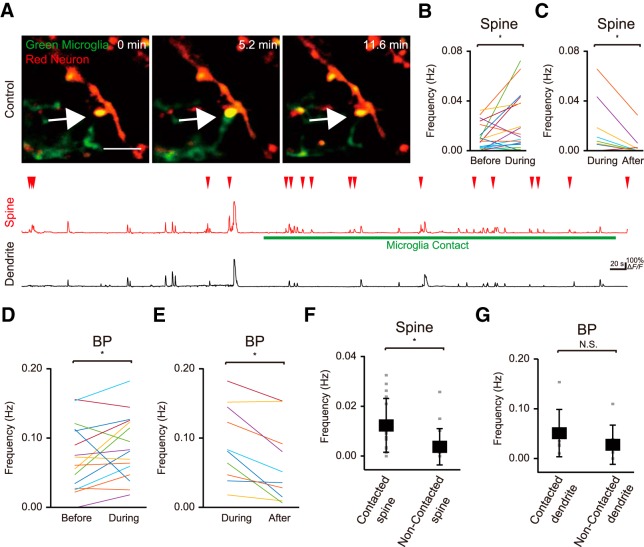
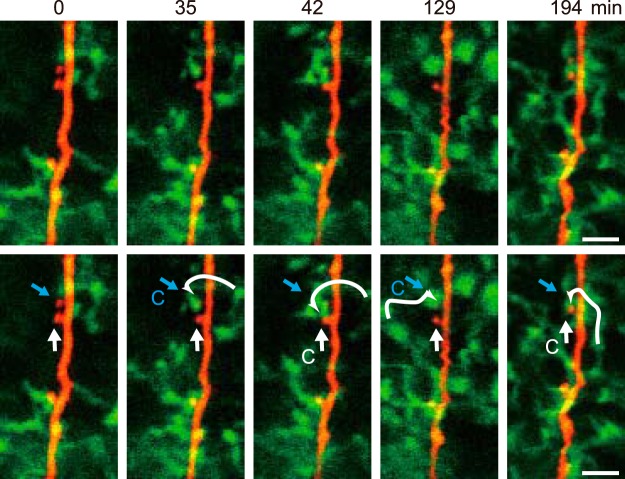
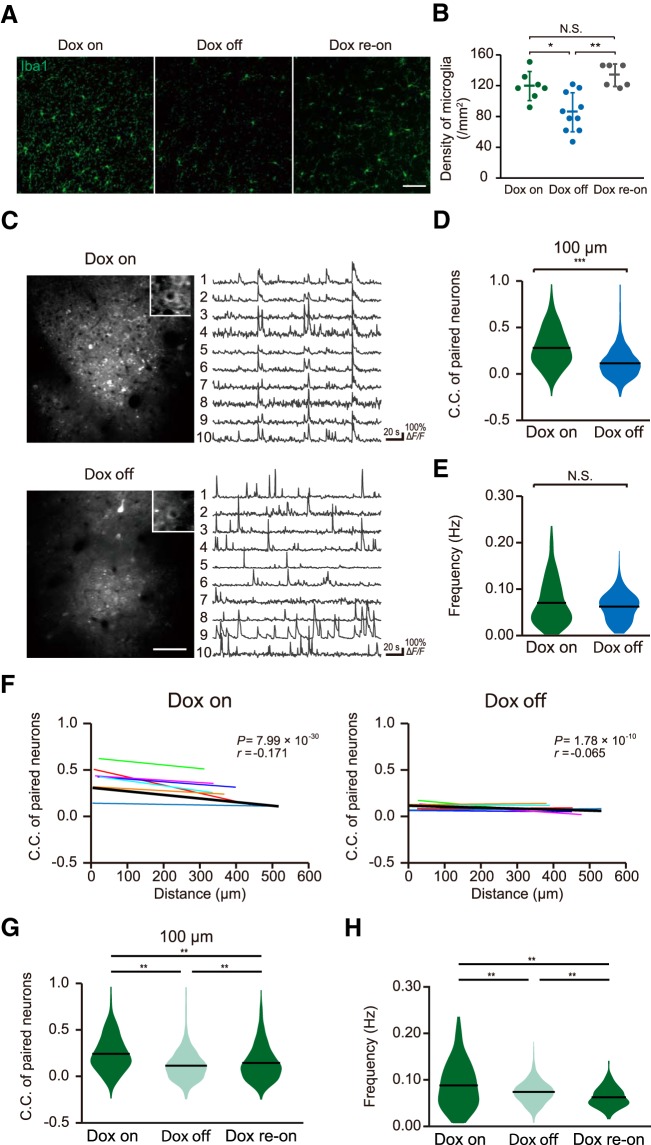
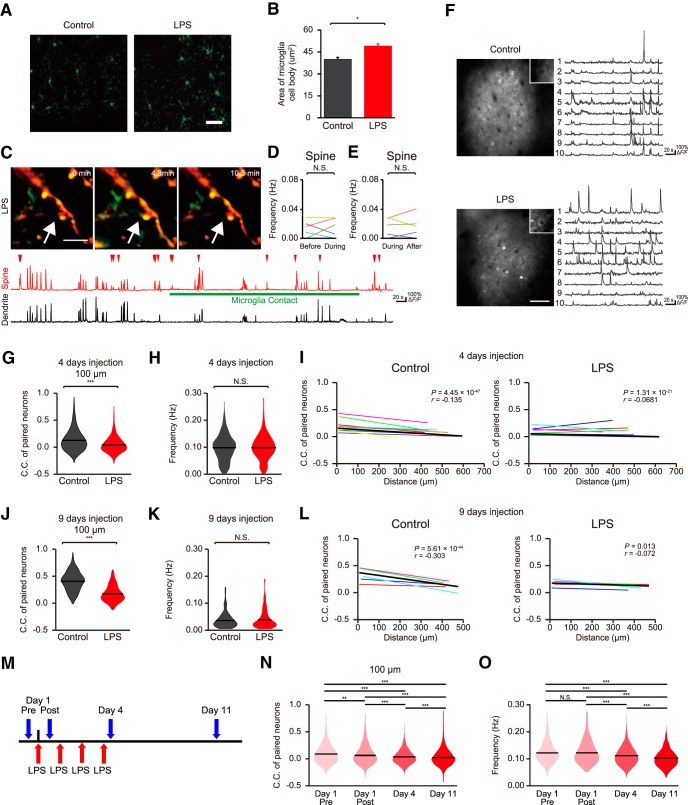
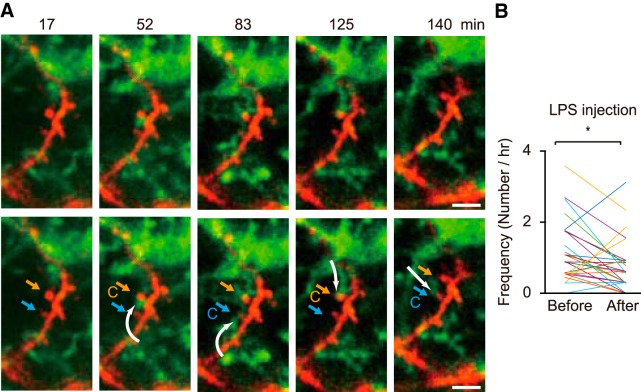
Microglia are highly motile immunoreactive cells that play integral roles in the response to brain infection and damage, and in the progression of various neurological diseases. During development, microglia also help sculpt neural circuits, via both promoting synapse formation and by targeting specific synapses for elimination and phagocytosis. Microglia are also active surveyors of neural circuits in the mature, healthy brain, although the functional consequences of such microglia-neuron contacts under these conditions is unclear. Using in vivo imaging of neurons and microglia in awake mice, we report here the functional consequences of microglia-synapse contacts. Direct contact between a microglial process and a single synapse results in a specific increase in the activity of that contacted synapse, and a corresponding increase in back-propagating action potentials along the parent dendrite. This increase in activity is not seen for microglia-synapse contacts when microglia are activated by chronic lipopolysaccharide (LPS) treatment. To probe how this microglia-synapse contact affects neural circuits, we imaged across larger populations of motor cortical neurons. When microglia were again activated by LPS (or partially ablated), there was a decrease in the extent to which neuronal activity was synchronized. Together, our results demonstrate that interactions between physiological or resting microglia and synapses in the mature, healthy brain leads to an increase in neuronal activity and thereby helps to synchronize local populations of neurons. Our novel findings provide a plausible physical basis for understanding how alterations in immune status may impact on neural circuit plasticity and on cognitive behaviors such as learning.
Keywords: calcium imaging; in vivo; microglia; synapse; two photon.
Figures
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources