

Tissue engineering the cancer microenvironment-challenges and opportunities
- PMID: 30406572
- PMCID: PMC6297082
- DOI: 10.1007/s12551-018-0466-8
Tissue engineering the cancer microenvironment-challenges and opportunities
Abstract
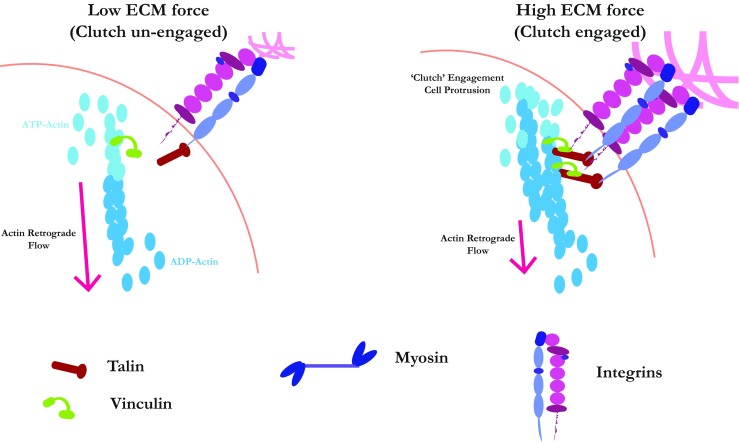
Mechanosensing is increasingly recognised as important for tumour progression. Tumours become stiff and the forces that normally balance in the healthy organism break down and become imbalanced, leading to increases in migration, invasion and metastatic dissemination. Here, we review recent advances in our understanding of how extracellular matrix properties, such as stiffness, viscoelasticity and architecture control cell behaviour. In addition, we discuss how the tumour microenvironment can be modelled in vitro, capturing these mechanical aspects, to better understand and develop therapies against tumour spread. We argue that by gaining a better understanding of the microenvironment and the mechanical forces that govern tumour dynamics, we can make advances in combatting cancer dormancy, recurrence and metastasis.
Keywords: Adhesion; Cancer microenvironment; Cell migration; Cytoskeleton; Extracellular matrix; Hydrogels; Mechanosensing; Motility.
Conflict of interest statement
Conflict of interest
Vassilis Papalazarou declares that he has no conflict of interest. Manuel Salmeron-Sanchez declares that he has no conflict of interest. Laura M. Machesky declares that she has no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Figures




References
-
- Ahmadzadeh H, Webster MR, Behera R, Jimenez Valencia AM, Wirtz D, Weeraratna AT, Shenoy VB. Modeling the two-way feedback between contractility and matrix realignment reveals a nonlinear mode of cancer cell invasion. Proc Natl Acad Sci U S A. 2017;114:E1617–E1626. doi: 10.1073/pnas.1617037114. - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
