

Hybridisation-based target enrichment of phenology genes to dissect the genetic basis of yield and adaptation in barley
- PMID: 30407713
- PMCID: PMC6587706
- DOI: 10.1111/pbi.13029
Hybridisation-based target enrichment of phenology genes to dissect the genetic basis of yield and adaptation in barley
Abstract
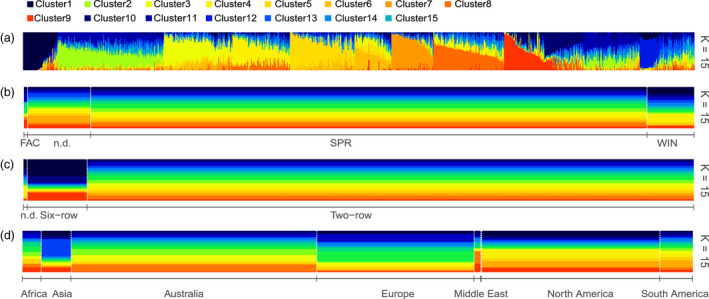
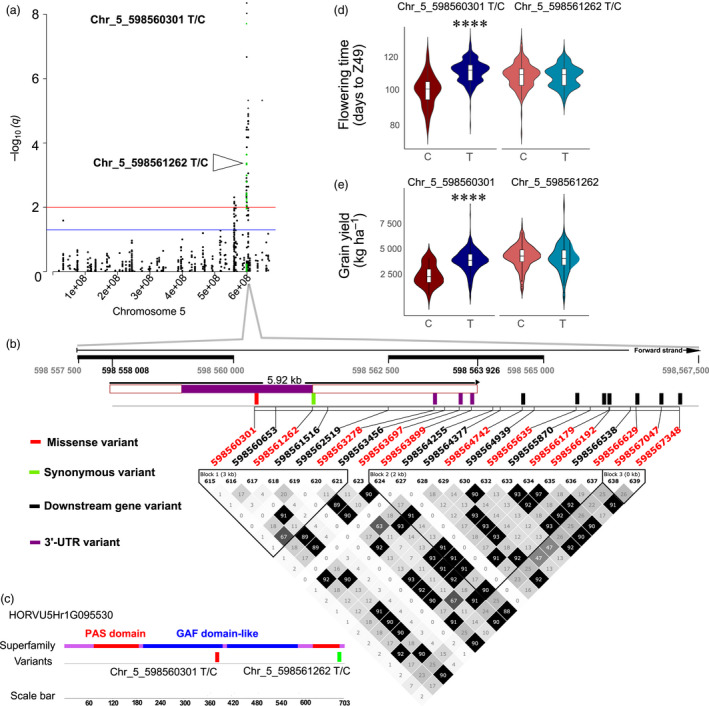
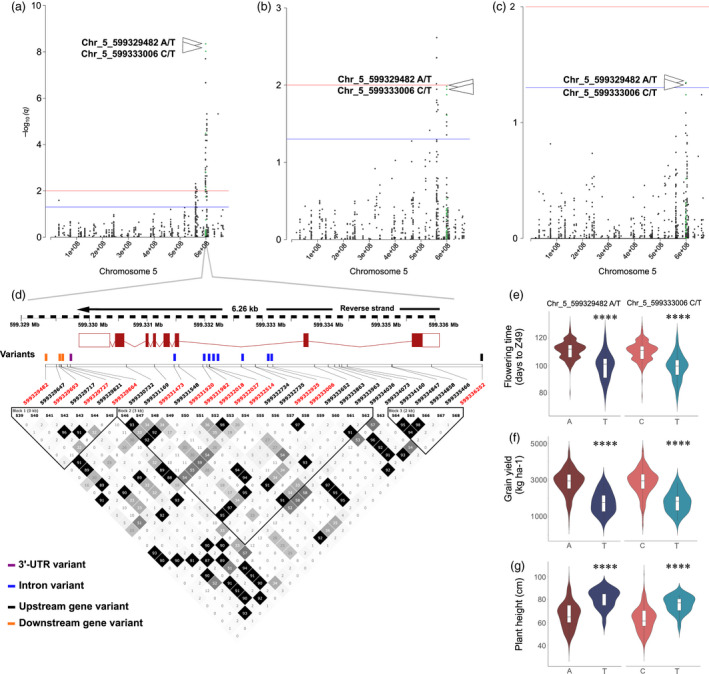
Barley (Hordeum vulgare L.) is a major cereal grain widely used for livestock feed, brewing malts and human food. Grain yield is the most important breeding target for genetic improvement and largely depends on optimal timing of flowering. Little is known about the allelic diversity of genes that underlie flowering time in domesticated barley, the genetic changes that have occurred during breeding, and their impact on yield and adaptation. Here, we report a comprehensive genomic assessment of a worldwide collection of 895 barley accessions based on the targeted resequencing of phenology genes. A versatile target-capture method was used to detect genome-wide polymorphisms in a panel of 174 flowering time-related genes, chosen based on prior knowledge from barley, rice and Arabidopsis thaliana. Association studies identified novel polymorphisms that accounted for observed phenotypic variation in phenology and grain yield, and explained improvements in adaptation as a result of historical breeding of Australian barley cultivars. We found that 50% of genetic variants associated with grain yield, and 67% of the plant height variation was also associated with phenology. The precise identification of favourable alleles provides a genomic basis to improve barley yield traits and to enhance adaptation for specific production areas.
Keywords: Hordeum vulgare; association mapping; flowering time; grain yield; next-generation sequencing; target capture.
© 2018 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
DM is affiliated with InterGrain Pty Ltd WA. PT is affiliated with Australian Grain Technologies Pty Ltd (AGT), SA. There are no financial relationships or competing interests on the part of any author that could potentially bias the findings reported in the manuscript.
Figures
Similar articles
-
Gene-set association and epistatic analyses reveal complex gene interaction networks affecting flowering time in a worldwide barley collection.J Exp Bot. 2019 Oct 24;70(20):5603-5616. doi: 10.1093/jxb/erz332. J Exp Bot. 2019. PMID: 31504706 Free PMC article.
-
Targeted enrichment by solution-based hybrid capture to identify genetic sequence variants in barley.Sci Data. 2019 Apr 1;6(1):12. doi: 10.1038/s41597-019-0011-z. Sci Data. 2019. PMID: 30931948 Free PMC article.
-
Modelling the genetic architecture of flowering time control in barley through nested association mapping.BMC Genomics. 2015 Apr 12;16(1):290. doi: 10.1186/s12864-015-1459-7. BMC Genomics. 2015. PMID: 25887319 Free PMC article.
-
Major flowering time genes of barley: allelic diversity, effects, and comparison with wheat.Theor Appl Genet. 2021 Jul;134(7):1867-1897. doi: 10.1007/s00122-021-03824-z. Epub 2021 May 9. Theor Appl Genet. 2021. PMID: 33969431 Free PMC article. Review.
-
Genetic Architecture of Flowering Phenology in Cereals and Opportunities for Crop Improvement.Front Plant Sci. 2016 Dec 19;7:1906. doi: 10.3389/fpls.2016.01906. eCollection 2016. Front Plant Sci. 2016. PMID: 28066466 Free PMC article. Review.
Cited by
-
New semi-dwarfing alleles with increased coleoptile length by gene editing of gibberellin 3-oxidase 1 using CRISPR-Cas9 in barley (Hordeum vulgare L.).Plant Biotechnol J. 2023 Apr;21(4):806-818. doi: 10.1111/pbi.13998. Epub 2023 Feb 8. Plant Biotechnol J. 2023. PMID: 36587283 Free PMC article.
-
Multi-locus genome-wide association studies reveal novel alleles for flowering time under vernalisation and extended photoperiod in a barley MAGIC population.Theor Appl Genet. 2022 Sep;135(9):3087-3102. doi: 10.1007/s00122-022-04169-x. Epub 2022 Jul 25. Theor Appl Genet. 2022. PMID: 35879467 Free PMC article.
-
Highly efficient and genotype-independent barley gene editing based on anther culture.Plant Commun. 2020 Jun 5;2(2):100082. doi: 10.1016/j.xplc.2020.100082. eCollection 2021 Mar 8. Plant Commun. 2020. PMID: 33898972 Free PMC article.
-
Genetic dissection of the interactions between semi-dwarfing genes sdw1 and ari-e and their effects on agronomic traits in a barley MAGIC population.Mol Breed. 2020 Jun 23;40(7):64. doi: 10.1007/s11032-020-01145-5. eCollection 2020 Jul. Mol Breed. 2020. PMID: 40255936 Free PMC article.
-
Genome-Wide Association Study and Identification of Candidate Genes for Nitrogen Use Efficiency in Barley (Hordeum vulgare L.).Front Plant Sci. 2020 Sep 4;11:571912. doi: 10.3389/fpls.2020.571912. eCollection 2020. Front Plant Sci. 2020. PMID: 33013994 Free PMC article.
References
-
- Abbo, S. , Lev‐Yadun, S. and Gopher, A. (2010) Agricultural origins: centers and noncenters; a Near Eastern reappraisal. Crit. Rev. Plant Sci. 29, 317–328.
-
- Andres, F. and Coupland, G. (2012) The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 13, 627–639. - PubMed
-
- Araus, J.L. and Cairns, J.E. (2014) Field high‐throughput phenotyping: the new crop breeding frontier. Trends Plant Sci. 19, 52–61. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
