

Sex-specific airway hyperreactivity and sex-specific transcriptome remodeling in neonatal piglets challenged with intra-airway acid
- PMID: 30407862
- PMCID: PMC6383496
- DOI: 10.1152/ajplung.00417.2018
Sex-specific airway hyperreactivity and sex-specific transcriptome remodeling in neonatal piglets challenged with intra-airway acid
Abstract
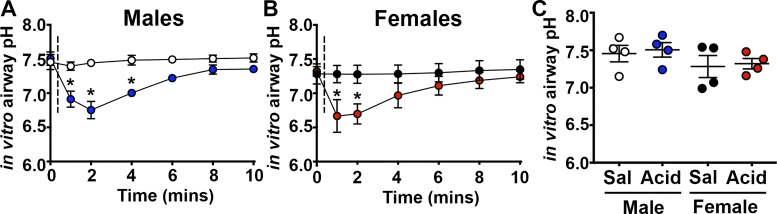
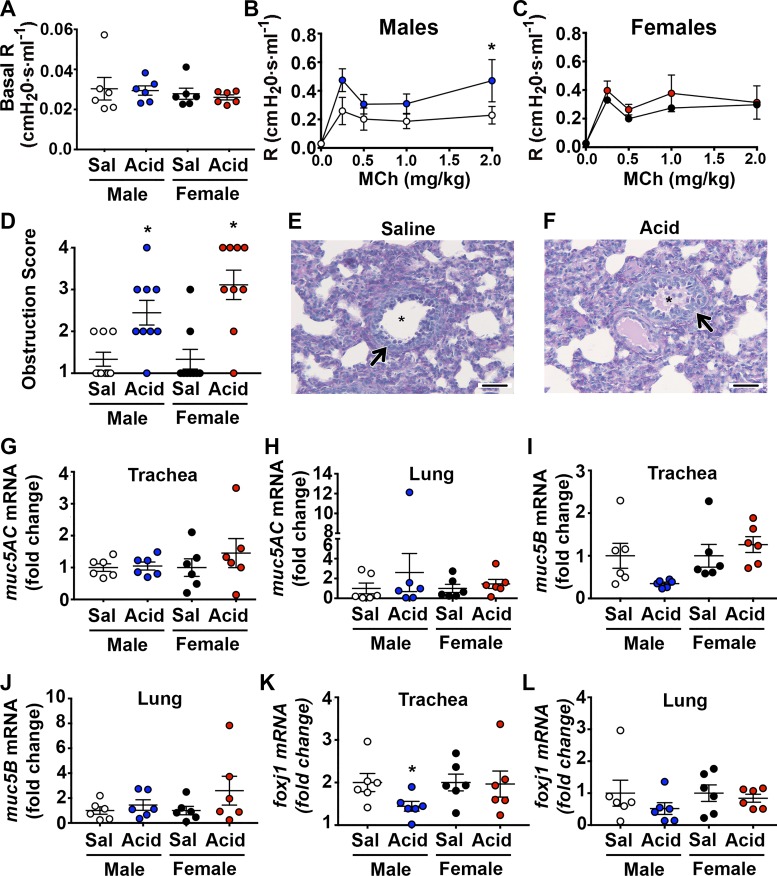
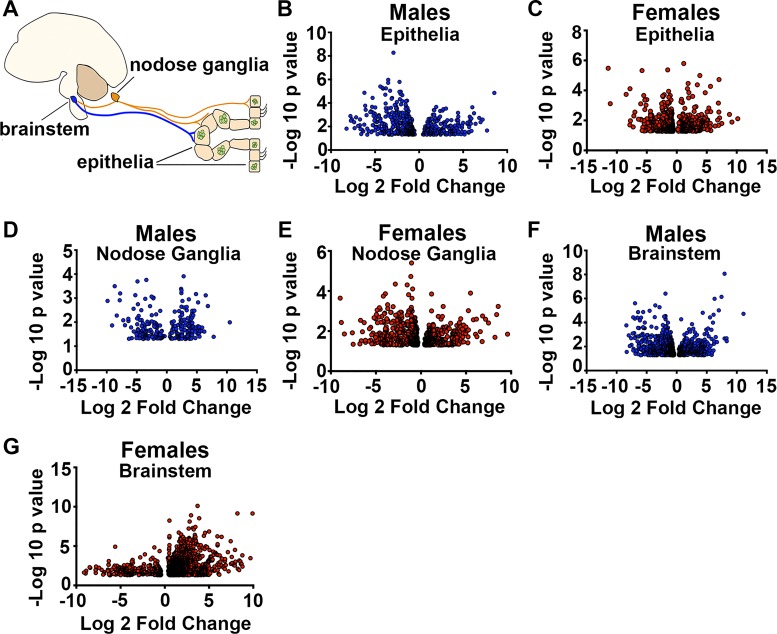
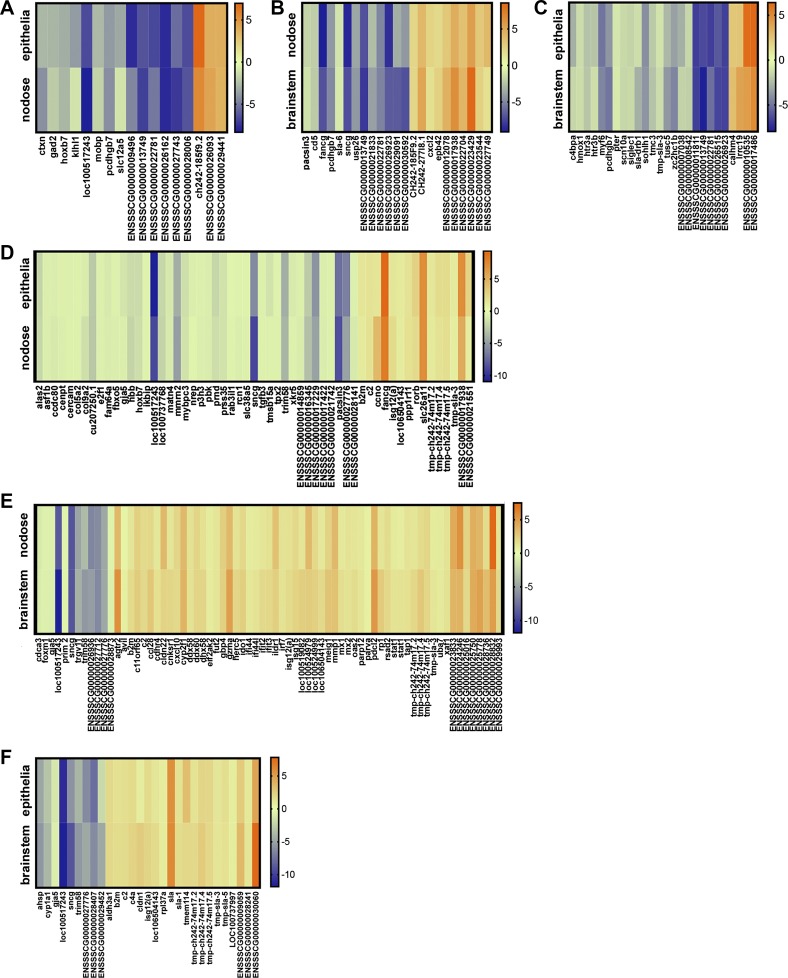
Acute airway acidification is a potent stimulus of sensory nerves and occurs commonly with gastroesophageal reflux disease, cystic fibrosis, and asthma. In infants and adults, airway acidification can acutely precipitate asthma-like symptoms, and treatment-resistant asthma can be associated with gastroesophageal reflux disease. Airway protective behaviors, such as mucus secretion and airway smooth muscle contraction, are often exaggerated in asthma. These behaviors are manifested through activation of neural circuits. In some populations, the neural response to acid might be particularly important. For example, the immune response in infants is relatively immature compared with adults. Infants also have a high frequency of gastroesophageal reflux. Thus, in the current study, we compared the transcriptomes of an airway-nervous system circuit (e.g., tracheal epithelia, nodose ganglia, and brain stem) in neonatal piglets challenged with intra-airway acid. We hypothesized that the identification of parallel changes in the transcriptomes of two neutrally connected tissues might reveal the circuit response, and, hence, molecules important for the manifestation of asthma-like features. Intra-airway acid induced airway hyperreactivity and airway obstruction in male piglets. In contrast, female piglets displayed airway obstruction without airway hyperreactivity. Pairwise comparisons revealed parallel changes in genes directly implicated in airway hyperreactivity ( scn10a) in male acid-challenged piglets, whereas acid-challenged females exhibited parallel changes in genes associated with mild asthma ( stat 1 and isg15). These findings reveal sex-specific responses to acute airway acidification and highlight distinct molecules within a neural circuit that might be critical for the manifestation of asthma-like symptoms in pediatric populations.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures


Similar articles
-
The presence of LPS in OVA inhalations affects airway inflammation and AHR but not remodeling in a rodent model of asthma.Am J Physiol Lung Cell Mol Physiol. 2012 Jul 1;303(1):L54-63. doi: 10.1152/ajplung.00208.2011. Epub 2012 Apr 20. Am J Physiol Lung Cell Mol Physiol. 2012. PMID: 22523281
-
Airway remodeling in asthma.Allergol Int. 2007 Dec;56(4):341-8. doi: 10.2332/allergolint.R-07-153. Epub 2007 Nov 1. Allergol Int. 2007. PMID: 17965577 Review.
-
Inhibition of airway inflammation, hyperresponsiveness and remodeling by soy isoflavone in a murine model of allergic asthma.Int Immunopharmacol. 2011 Aug;11(8):899-906. doi: 10.1016/j.intimp.2011.02.001. Epub 2011 Feb 26. Int Immunopharmacol. 2011. PMID: 21354484
-
Exchange protein directly activated by cAMP (Epac) protects against airway inflammation and airway remodeling in asthmatic mice.Respir Res. 2019 Dec 18;20(1):285. doi: 10.1186/s12931-019-1260-2. Respir Res. 2019. PMID: 31852500 Free PMC article.
-
Acid stress in the pathology of asthma.J Allergy Clin Immunol. 2004 Apr;113(4):610-9. doi: 10.1016/j.jaci.2003.12.034. J Allergy Clin Immunol. 2004. PMID: 15100663 Review.
Cited by
-
Airway cholinergic history modifies mucus secretion properties to subsequent cholinergic challenge in diminished chloride and bicarbonate conditions.Exp Physiol. 2020 Oct;105(10):1673-1683. doi: 10.1113/EP088900. Epub 2020 Aug 12. Exp Physiol. 2020. PMID: 32735372 Free PMC article.
-
Acid exposure disrupts mucus secretion and impairs mucociliary transport in neonatal piglet airways.Am J Physiol Lung Cell Mol Physiol. 2020 May 1;318(5):L873-L887. doi: 10.1152/ajplung.00025.2020. Epub 2020 Mar 11. Am J Physiol Lung Cell Mol Physiol. 2020. PMID: 32160007 Free PMC article.
-
Overexpression of Substance P in pig airways increases MUC5AC through an NF-kβ pathway.Physiol Rep. 2021 Feb;9(3):e14749. doi: 10.14814/phy2.14749. Physiol Rep. 2021. PMID: 33580593 Free PMC article.
-
Strategies for measuring airway mucus and mucins.Respir Res. 2019 Nov 21;20(1):261. doi: 10.1186/s12931-019-1239-z. Respir Res. 2019. PMID: 31752894 Free PMC article. Review.
-
Long-term culturing of porcine nodose ganglia.J Neurosci Methods. 2020 Feb 15;332:108546. doi: 10.1016/j.jneumeth.2019.108546. Epub 2019 Dec 9. J Neurosci Methods. 2020. PMID: 31821820 Free PMC article.
References
-
- Adam RJ, Michalski AS, Bauer C, Abou Alaiwa MH, Gross TJ, Awadalla MS, Bouzek DC, Gansemer ND, Taft PJ, Hoegger MJ, Diwakar A, Ochs M, Reinhardt JM, Hoffman EA, Beichel RR, Meyerholz DK, Stoltz DA. Air trapping and airflow obstruction in newborn cystic fibrosis piglets. Am J Respir Crit Care Med 188: 1434–1441, 2013. doi:10.1164/rccm.201307-1268OC. - DOI - PMC - PubMed
-
- Al-Alwan LA, Chang Y, Mogas A, Halayko AJ, Baglole CJ, Martin JG, Rousseau S, Eidelman DH, Hamid Q. Differential roles of CXCL2 and CXCL3 and their receptors in regulating normal and asthmatic airway smooth muscle cell migration. J Immunol 191: 2731–2741, 2013. doi:10.4049/jimmunol.1203421. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous